Methionine
 Canonical form of methionine | |
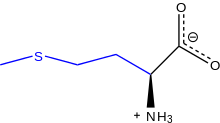 Resonance line-angle diagram of L-methionine at physiological pH | |
| Names | |
|---|---|
IUPAC name Methionine | |
| Other names 2-amino-4-(methylthio)butanoic acid | |
| Identifiers | |
CAS Number |
|
3D model (JSmol) |
|
| Abbreviations | Met, M |
ChEBI |
|
ChEMBL |
|
ChemSpider |
|
ECHA InfoCard | 100.000.393 |
EC Number | 200-432-1 |
KEGG |
|
PubChem CID |
|
UNII |
|
InChI
| |
SMILES
| |
| Properties[2] | |
Chemical formula | C5H11NO2S |
Molar mass | 149.21 g·mol−1 |
| Appearance | White crystalline powder |
Density | 1.340 g/cm3 |
Melting point | 281 °C (538 °F; 554 K) decomposes |
Solubility in water | Soluble |
Acidity (pKa) | 2.28 (carboxyl), 9.21 (amino)[1] |
| Pharmacology | |
ATC code | V03AB26 (WHO) QA05BA90 (WHO), QG04BA90 (WHO) |
Supplementary data page | |
Structure and properties | Refractive index (n), Dielectric constant (εr), etc. |
Thermodynamic data | Phase behaviour solid–liquid–gas |
Spectral data | UV, IR, NMR, MS |
Except where otherwise noted, data are given for materials in their standard state (at 25 °C [77 °F], 100 kPa). | |
Infobox references | |
Methionine (symbol Met or M)[3] (/mɪˈθaɪəniːn/)[4] is an essential amino acid in humans. As the substrate for other amino acids such as cysteine and taurine, versatile compounds such as SAM-e, and the important antioxidant glutathione, methionine plays a critical role in the metabolism and health of many species, including humans. It is encoded by the codon AUG.
Methionine is also an important part of angiogenesis, the growth of new blood vessels. Supplementation may benefit those suffering from copper poisoning.[5] Overconsumption of methionine, the methyl group donor in DNA methylation, is related to cancer growth in a number of studies.[6][7] Methionine was first isolated in 1921 by John Howard Mueller.[8]
Contents
1 Biomedical details
2 A proteinogenic amino acid
2.1 Encoding
3 Derivatives
3.1 S-adenosyl-methionine
4 Biosynthesis
4.1 Trans-sulfurylation pathway
5 Other biochemical pathways
5.1 Catabolism
5.2 Regeneration
5.3 Reverse-transulfurylation pathway: conversion to cysteine
5.4 Ethylene synthesis
6 Chemical synthesis
7 Human nutrition
7.1 Requirements
7.2 Dietary sources
8 Restriction
9 Health
10 Other uses
11 See also
12 References
13 External links
Biomedical details
Methionine (abbreviated as Met or M; encoded by the codon AUG) is an α-amino acid that is used in the biosynthesis of proteins. It contains an α-amino group (which is in the protonated −NH3+ form under biological conditions), an α-carboxylic acid group (which is in the deprotonated −COO− form under biological conditions), and an S-methyl thioether side chain, classifying it as a nonpolar, aliphatic amino acid.
Methionine is coded for by the initiation codon, meaning it indicates the start of the coding region and is the first amino acid produced in a nascent polypeptide during mRNA translation.[9]
A proteinogenic amino acid
Together with cysteine, methionine is one of two sulfur-containing proteinogenic amino acids. Excluding the few exceptions where methionine may act as a redox sensor (e.g.,[10]), methionine residues do not have a catalytic role.[11] This is in contrast to cysteine residues, where the thiol group has a catalytic role in many proteins.[11] The thioether does however have a minor structural role due to the stability effect of S/π interactions between the side chain sulfur atom and aromatic amino acids in one-third of all known protein structures.[11] This lack of a strong role is reflected in experiments where little effect is seen in proteins where methionine is replaced by norleucine, a straight hydrocarbon sidechain amino acid which lacks the thioether.[12]
It has been conjectured that norleucine was present in early versions of the genetic code, but methionine intruded into the final version of the genetic code due to the fact it is used in the cofactor S-adenosyl methionine (SAM).[13] This situation is not unique and may have occurred with ornithine and arginine.[14]
Encoding
Methionine is one of only two amino acids encoded by a single codon (AUG) in the standard genetic code (tryptophan, encoded by UGG, is the other). In reflection to the evolutionary origin of its codon, the other AUN codons encode isoleucine, which is also a hydrophobic amino acid. In the mitochondrial genome of several organisms, including metazoa and yeast, the codon AUA also encodes for methionine. In the standard genetic code AUA codes for isoleucine and the respective tRNA (ileX in Escherichia coli) uses the unusual base lysidine (bacteria) or agmatine (archaea) to discriminate against AUG.[15][16]
The methionine codon AUG is also the most common start codon. A "Start" codon is message for a ribosome that signals the initiation of protein translation from mRNA when the AUG codon is in a Kozak consensus sequence. As a consequence, methionine is often incorporated into the N-terminal position of proteins in eukaryotes and archaea during translation, although it can be removed by post-translational modification. In bacteria, the derivative N-formylmethionine is used as the initial amino acid.
Derivatives
S-adenosyl-methionine
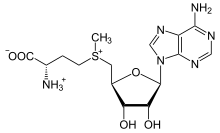
S-Adenosyl-methionine is a cofactor derived from methionine.
The methionine-derivative S-adenosyl methionine (SAM) is a cofactor that serves mainly as a methyl donor. SAM is composed of an adenosyl molecule (via 5' carbon) attached to the sulfur of methionine, therefore making it a sulfonium cation (i.e., three substituents and positive charge). The sulfur acts as a soft Lewis acid (i.e., donor/electrophile) which allows the S-methyl group to be transferred to an oxygen, nitrogen, or aromatic system, often with the aid of other cofactors such as cobalamin (vitamin B12 in humans). Some enzymes use SAM to initiate a radical reaction; these are called radical SAM enzymes.
As a result of the transfer of the methyl group, S-adenosyl-homocysteine is obtained. In bacteria, this is either regenerated by methylation or is salvaged by removing the adenine and the homocysteine, leaving the compound dihydroxypentandione to spontaneously convert into autoinducer-2, which is excreted as a waste product / quorum signal.
Biosynthesis

Methionine biosynthesis
As an essential amino acid, methionine is not synthesized de novo in humans and other animals, which must ingest methionine or methionine-containing proteins. In plants and microorganisms, methionine biosynthesis belongs to the aspartate family, along with threonine and lysine (via diaminopimelate, but not via α-aminoadipate). The main backbone is derived from aspartic acid, while the sulfur may come from cysteine, methanethiol, or hydrogen sulfide.[11]
- First, aspartic acid is converted via β-aspartyl-semialdehyde into homoserine by two reduction steps of the terminal carboxyl group (homoserine has therefore a γ-hydroxyl, hence the homo- series). The intermediate aspartate-semialdehyde is the branching point with the lysine biosynthetic pathway, where it is instead condensed with pyruvate. Homoserine is the branching point with the threonine pathway, where instead it is isomerised after activating the terminal hydroxyl with phosphate (also used for methionine biosynthesis in plants).[11]
- Homoserine is then activated with a phosphate, succinyl or an acetyl group on the hydroxyl.
- In plants and possibly in some bacteria,[11] phosphate is used. This step is shared with threonine biosynthesis.[11]
- In most organisms, an acetyl group is used to activate the homoserine. This can be catalysed in bacteria by an enzyme encoded by metX or metA (not homologues).[11]
- In enterobacteria and a limited number of other organisms, succinate is used. The enzyme that catalyses the reaction is MetA and the specificity for acetyl-CoA and succinyl-CoA is dictated by a single reside.[11] The physiological basis for the preference of acetyl-CoA or succinyl-CoA is unknown, but such alternative routes are present in some other pathways (e.g. lysine biosynthesis and arginine biosynthesis).
- In plants and possibly in some bacteria,[11] phosphate is used. This step is shared with threonine biosynthesis.[11]
- The hydroxyl activating group is then replaced with cysteine, methanethiol, or hydrogen sulfide. A replacement reaction is technically a γ-elimination followed by a variant of a Michael addition. All the enzymes involved are homologues and members of the Cys/Met metabolism PLP-dependent enzyme family, which is a subset of the PLP-dependent fold type I clade. They utilise the cofactor PLP (pyridoxal phosphate), which functions by stabilising carbanion intermediates.[11]
- If it reacts with cysteine, it produces cystathionine, which is cleaved to yield homocysteine. The enzymes involved are cystathionine-γ-synthase (encoded by metB in bacteria) and cystathionine-β-lyase (metC). Cystathionine is bound differently in the two enzymes allowing β or γ reactions to occur.[11]
- If it reacts with free hydrogen sulfide, it produces homocysteine. This is catalysed by O-acetylhomoserine aminocarboxypropyltransferase (formerly known as O-acetylhomoserine (thiol)-lyase. It is encoded by either metY or metZ in bacteria.[11]
- If it reacts with methanethiol, it produces methionine directly. Methanethiol is a byproduct of catabolic pathway of certain compounds, therefore this route is more uncommon.[11]
- If it reacts with cysteine, it produces cystathionine, which is cleaved to yield homocysteine. The enzymes involved are cystathionine-γ-synthase (encoded by metB in bacteria) and cystathionine-β-lyase (metC). Cystathionine is bound differently in the two enzymes allowing β or γ reactions to occur.[11]
- If homocysteine is produced, the thiol group is methylated, yielding methionine. Two methionine synthases are known; one is cobalamin (vitamin B12) dependent and one is independent.[11]
The pathway using cysteine is called the "transsulfuration pathway", while the pathway using hydrogen sulfide (or methanethiol) is called "direct-sulfurylation pathway".
Cysteine is similarly produced, namely it can be made from an activated serine and either from homocysteine ("reverse trans-sulfurylation route") or from hydrogen sulfide ("direct sulfurylation route"); the activated serine is generally O-acetyl-serine (via CysK or CysM in E. coli), but in Aeropyrum pernix and some other archaea O-phosphoserine is used.[17] CysK and CysM are homologues, but belong to the PLP fold type III clade.
Trans-sulfurylation pathway
Enzymes involved in the E. coli trans-sulfurylation route of methionine biosynthesis:
- Aspartokinase
- Aspartate-semialdehyde dehydrogenase
- Homoserine dehydrogenase
- Homoserine O-transsuccinylase
- Cystathionine-γ-synthase
- Cystathionine-β-lyase
Methionine synthase (in mammals, this step is performed by homocysteine methyltransferase or betaine—homocysteine S-methyltransferase.)
Other biochemical pathways

Fates of methionine
Although mammals cannot synthesize methionine, they can still use it in a variety of biochemical pathways:
Catabolism
Methionine is converted to S-adenosylmethionine (SAM) by (1) methionine adenosyltransferase.
SAM serves as a methyl-donor in many (2) methyltransferase reactions, and is converted to S-adenosylhomocysteine (SAH).
(3) Adenosylhomocysteinase converts SAH to homocysteine.
Homocysteine can be used to regenerate methionine or to form cysteine.
Regeneration
Methionine can be regenerated from homocysteine via (4) methionine synthase in a reaction that requires vitamin B12 as a cofactor.
Homocysteine can also be remethylated using glycine betaine (NNN-trimethyl glycine, TMG) to methionine via the enzyme betaine-homocysteine methyltransferase (E.C.2.1.1.5, BHMT). BHMT makes up to 1.5% of all the soluble protein of the liver, and recent evidence suggests that it may have a greater influence on methionine and homocysteine homeostasis than methionine synthase.
Reverse-transulfurylation pathway: conversion to cysteine
Homocysteine can be converted to cysteine.
- (5) Cystathionine-β-synthase (an enzyme which requires the active form of vitamin B6, pyridoxal phosphate) combines homocysteine and serine to produce cystathionine. Instead of degrading cystathionine via cystathionine-β-lyase, as in the biosynthetic pathway, cystathionine is broken down to cysteine and α-ketobutyrate via (6) cystathionine-γ-lyase.
- (7) The enzyme α-ketoacid dehydrogenase converts α-ketobutyrate to propionyl-CoA, which is metabolized to succinyl-CoA in a three-step process (see propionyl-CoA for pathway).
Ethylene synthesis
This amino acid is also used by plants for synthesis of ethylene. The process is known as the Yang cycle or the methionine cycle.

The Yang cycle
Chemical synthesis
Racemic methionine can be synthesized from diethyl sodium phthalimidomalonate by alkylation with chloroethylmethylsulfide (ClCH2CH2SCH3) followed by hydrolysis and decarboxylation.[18]
Human nutrition
Requirements
The Food and Nutrition Board of the U.S. Institute of Medicine set Recommended Dietary Allowances (RDAs) for essential amino acids in 2002. For methionine combined with cysteine, for adults 19 years and older, 19 mg/kg body weight/day.[19]
Dietary sources
| Food | g/100g |
|---|---|
Egg, white, dried, powder, glucose reduced | 3.204 |
Sesame seeds flour (low fat) | 1.656 |
Egg, whole, dried | 1.477 |
Cheese, Parmesan, shredded | 1.114 |
| Brazil nuts | 1.008 |
| Soy protein concentrate | 0.814 |
Chicken, broilers or fryers, roasted | 0.801 |
Fish, tuna, light, canned in water, drained solids | 0.755 |
Beef, cured, dried | 0.749 |
| Bacon | 0.593 |
Beef, ground, 95% lean meat / 5% fat, raw | 0.565 |
Pork, ground, 96% lean / 4% fat, raw | 0.564 |
| Wheat germ | 0.456 |
| Oat | 0.312 |
| Peanuts | 0.309 |
| Chickpea | 0.253 |
Corn, yellow | 0.197 |
| Almonds | 0.151 |
| Beans, pinto, cooked | 0.117 |
Lentils, cooked | 0.077 |
Rice, brown, medium-grain, cooked | 0.052 |
High levels of methionine can be found in eggs, meat, and fish; sesame seeds, Brazil nuts, and some other plant seeds; and cereal grains. Most fruits and vegetables contain very little. Most legumes, though protein dense, are low in methionine. Proteins without adequate methionine are not considered to be complete proteins.[21] For that reason, racemic methionine is sometimes added as an ingredient to pet foods.[22]
Restriction
Some scientific evidence indicates restricting methionine consumption can increase lifespans in fruit flies.[23]
A 2005 study showed methionine restriction without energy restriction extends mouse lifespans.[24] This extension requires intact growth hormone signaling, as animals without intact growth-hormone signaling do not have a further increase in lifespan when methionine restricted.[25] The metabolic response to methionine restriction is also altered in mouse growth hormone signaling mutants.[26]
A study published in Nature showed adding just the essential amino acid methionine to the diet of fruit flies under dietary restriction, including restriction of essential amino acids (EAAs), restored fertility without reducing the longer lifespans that are typical of dietary restriction, leading the researchers to determine that methionine “acts in combination with one or more other EAAs to shorten lifespan.”[23][27][28] Restoring methionine to the diet of mice on a dietary restriction regimen blocks many acute benefits of dietary restriction, a process that may be mediated by increased production of hydrogen sulfide.[29]
Several studies showed that methionine restriction also inhibits aging-related disease processes in mice[30][31] and inhibits colon carcinogenesis in rats.[32] In humans, methionine restriction through dietary modification could be achieved through a plant-based diet.[33]
A 2009 study on rats showed "methionine supplementation in the diet specifically increases mitochondrial ROS production and mitochondrial DNA oxidative damage in rat liver mitochondria offering a plausible mechanism for its hepatotoxicity".[34]
However, since methionine is an essential amino acid, it cannot be entirely removed from animals' diets without disease or death occurring over time.[citation needed] For example, rats fed a diet without methionine and choline developed steatohepatitis (fatty liver) and anemia, and lost two-thirds of their body weight over 5 weeks. Administration of methionine ameliorated the pathological consequences of methionine deprivation.[35] Short-term removal of only methionine from the diet can reverse diet-induced obesity and promotes insulin sensitivity in mice.[36]
Health
Loss of methionine has been linked to senile greying of hair. Its lack leads to a buildup of hydrogen peroxide in hair follicles, a reduction in tyrosinase effectiveness, and a gradual loss of hair color.[37]
Methionine is an intermediate in the biosynthesis of cysteine, carnitine, taurine, lecithin, phosphatidylcholine, and other phospholipids. Improper conversion of methionine can lead to atherosclerosis[38] due to accumulation of homocysteine.
Methionine might also be essential to reversing damaging methylation of glucocorticoid receptors caused by repeated stress exposures, with implications for depression.[39]
Other uses
DL-Methionine is sometimes given as a supplement to dogs; It helps to reduce the chances of stones in dogs. Methionine is also known to increase the urinary excretion of quinidine by acidifying the urine. Aminoglycoside antibiotics used to treat urinary tract infections work best in alkaline conditions, and urinary acidification from using methionine can reduce its effectiveness. If a dog is on a diet that acidifies the urine, methionine should not be used.[40]
Methionine is allowed as a supplement to organic poultry feed under the US certified organic program.[41]
Methionine can be used as a nontoxic pesticide option against giant swallowtail caterpillars, which are a serious pest to orange crops.[42]
See also
- Allantoin
- Formylmethionine
- Methionine oxidation
- Paracetamol poisoning
- Photoreactive methionine
References
^ Dawson RM, Elliott DC, Elliott WH, Jones KM (1959). Data for Biochemical Research. Oxford: Clarendon Press..mw-parser-output cite.citation{font-style:inherit}.mw-parser-output q{quotes:"""""""'""'"}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
^ Weast, Robert C., ed. (1981). CRC Handbook of Chemistry and Physics (62nd ed.). Boca Raton, FL: CRC Press. p. C-374. ISBN 0-8493-0462-8..
^ "Nomenclature and Symbolism for Amino Acids and Peptides". IUPAC-IUB Joint Commission on Biochemical Nomenclature. 1983. Archived from the original on 9 October 2008. Retrieved 5 March 2018.
^ "Methionine". Oxford University Press.
^ "Methionine". WebMD.
^ Cavuoto P, Fenech MF (2012). "A review of methionine dependency and the role of methionine restriction in cancer growth control and life-span extension". Cancer Treatment Reviews. 38 (6): 726–36. doi:10.1016/j.ctrv.2012.01.004. PMID 22342103.
^ Cellarier E, Durando X, Vasson MP, Farges MC, Demiden A, Maurizis JC, Madelmont JC, Chollet P (2003). "Methionine dependency and cancer treatment". Cancer Treatment Reviews. 29 (6): 489–99. doi:10.1016/S0305-7372(03)00118-X. PMID 14585259.
^ Pappenheimer AM (1987). "A Biographical Memoir of John Howard Mueller" (PDF). Washington D.C.: National Academy of Sciences.
^ Guedes RL, Prosdocimi F, Fernandes GR, Moura LK, Ribeiro HA, Ortega JM (December 2011). "Amino acids biosynthesis and nitrogen assimilation pathways: a great genomic deletion during eukaryotes evolution". BMC Genomics. 12 Suppl 4: S2. doi:10.1186/1471-2164-12-S4-S2. PMC 3287585. PMID 22369087.
^ Bigelow DJ, Squier TC (January 2005). "Redox modulation of cellular signaling and metabolism through reversible oxidation of methionine sensors in calcium regulatory proteins". Biochimica et Biophysica Acta (Submitted manuscript). 1703 (2): 121–34. doi:10.1016/j.bbapap.2004.09.012. PMID 15680220.
^ abcdefghijklmn Ferla MP, Patrick WM (August 2014). "Bacterial methionine biosynthesis". Microbiology. 160 (Pt 8): 1571–84. doi:10.1099/mic.0.077826-0. PMID 24939187.
^ Cirino PC, Tang Y, Takahashi K, Tirrell DA, Arnold FH (September 2003). "Global incorporation of norleucine in place of methionine in cytochrome P450 BM-3 heme domain increases peroxygenase activity". Biotechnology and Bioengineering. 83 (6): 729–34. doi:10.1002/bit.10718. PMID 12889037.
^ Alvarez-Carreño C, Becerra A, Lazcano A (October 2013). "Norvaline and norleucine may have been more abundant protein components during early stages of cell evolution". Origins of Life and Evolution of the Biosphere. 43 (4–5): 363–75. Bibcode:2013OLEB...43..363A. doi:10.1007/s11084-013-9344-3. PMID 24013929.
^ Jukes TH (August 1973). "Arginine as an evolutionary intruder into protein synthesis". Biochemical and Biophysical Research Communications. 53 (3): 709–14. doi:10.1016/0006-291x(73)90151-4. PMID 4731949.
^ Ikeuchi Y, Kimura S, Numata T, Nakamura D, Yokogawa T, Ogata T, Wada T, Suzuki T, Suzuki T (April 2010). "Agmatine-conjugated cytidine in a tRNA anticodon is essential for AUA decoding in archaea". Nature Chemical Biology. 6 (4): 277–82. doi:10.1038/nchembio.323. PMID 20139989.
^ Muramatsu T, Nishikawa K, Nemoto F, Kuchino Y, Nishimura S, Miyazawa T, Yokoyama S (November 1988). "Codon and amino-acid specificities of a transfer RNA are both converted by a single post-transcriptional modification". Nature. 336 (6195): 179–81. Bibcode:1988Natur.336..179M. doi:10.1038/336179a0. PMID 3054566.
^ Mino K, Ishikawa K (September 2003). "A novel O-phospho-L-serine sulfhydrylation reaction catalyzed by O-acetylserine sulfhydrylase from Aeropyrum pernix K1". FEBS Letters. 551 (1–3): 133–8. doi:10.1016/S0014-5793(03)00913-X. PMID 12965218.
^ Barger G, Weichselbaum TE (1934). "dl-Methionine". Organic Syntheses. 14: 58.; Collective Volume, 2, p. 384
^ Institute of Medicine (2002). "Protein and Amino Acids". Dietary Reference Intakes for Energy, Carbohydrates, Fiber, Fat, Fatty Acids, Cholesterol, Protein, and Amino Acids. Washington, DC: The National Academies Press. pp. 589–768.
^ "National Nutrient Database for Standard Reference". U.S. Department of Agriculture. Archived from the original on 2015-03-03. Retrieved 2009-09-07.
^ Finkelstein JD (May 1990). "Methionine metabolism in mammals". The Journal of Nutritional Biochemistry. 1 (5): 228–37. PMID 15539209.
^ Palika L (1996). The Consumer's Guide to Dog Food: What's in Dog Food, Why It's There and How to Choose the Best Food for Your Dog. New York: Howell Book House. ISBN 978-0-87605-467-3.
^ ab Grandison RC, Piper MD, Partridge L (December 2009). "Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila". Nature. 462 (7276): 1061–4. Bibcode:2009Natur.462.1061G. doi:10.1038/nature08619. PMC 2798000. PMID 19956092. Lay summary.
^ Miller RA, Buehner G, Chang Y, Harper JM, Sigler R, Smith-Wheelock M (June 2005). "Methionine-deficient diet extends mouse lifespan, slows immune and lens aging, alters glucose, T4, IGF-I and insulin levels, and increases hepatocyte MIF levels and stress resistance". Aging Cell. 4 (3): 119–25. doi:10.1111/j.1474-9726.2005.00152.x. PMID 15924568..
^ Brown-Borg, Holly M.; Rakoczy, Sharlene G.; Wonderlich, Joseph A.; Rojanathammanee, Lalida; Kopchick, John J.; Armstrong, Vanessa; Raasakka, Debbie (2014-12). "Growth hormone signaling is necessary for lifespan extension by dietary methionine". Aging Cell. 13 (6): 1019–1027. doi:10.1111/acel.12269. ISSN 1474-9726. PMC 4244257. PMID 25234161. Check date values in:|date=(help)
^ Brown-Borg, Holly M.; Rakoczy, Sharlene; Wonderlich, Joseph A.; Borg, Kurt E.; Rojanathammanee, Lalida (2018-4). "Metabolic adaptation of short-living growth hormone transgenic mice to methionine restriction and supplementation". Annals of the New York Academy of Sciences. 1418 (1): 118–136. doi:10.1111/nyas.13687. ISSN 1749-6632. PMID 29722030. Check date values in:|date=(help)
^ Grandison RC, Piper MD, Partridge L (December 2009). "Amino-acid imbalance explains extension of lifespan by dietary restriction in Drosophila". Nature. 462 (7276): 1061–4. Bibcode:2009Natur.462.1061G. doi:10.1038/nature08619. PMC 2798000. PMID 19956092.
^ "Amino acid recipe could be right for long life". Science News. December 2, 2009.
^ Hine C, Harputlugil E, Zhang Y, Ruckenstuhl C, Lee BC, Brace L, Longchamp A, Treviño-Villarreal JH, Mejia P, Ozaki CK, Wang R, Gladyshev VN, Madeo F, Mair WB, Mitchell JR (January 2015). "Endogenous hydrogen sulfide production is essential for dietary restriction benefits". Cell. 160 (1–2): 132–44. doi:10.1016/j.cell.2014.11.048. PMC 4297538. PMID 25542313.
^ Richie JP, Leutzinger Y, Parthasarathy S, Malloy V, Orentreich N, Zimmerman JA (December 1994). "Methionine restriction increases blood glutathione and longevity in F344 rats". FASEB Journal. 8 (15): 1302–7. PMID 8001743.
^ Sun L, Sadighi Akha AA, Miller RA, Harper JM (July 2009). "Life-span extension in mice by preweaning food restriction and by methionine restriction in middle age". The Journals of Gerontology. Series A, Biological Sciences and Medical Sciences. 64 (7): 711–22. doi:10.1093/gerona/glp051. PMC 2691799. PMID 19414512.
^ Komninou D, Leutzinger Y, Reddy BS, Richie JP (2006). "Methionine restriction inhibits colon carcinogenesis". Nutrition and Cancer. 54 (2): 202–8. doi:10.1207/s15327914nc5402_6. PMID 16898864.
^ Cavuoto P, Fenech MF (October 2012). "A review of methionine dependency and the role of methionine restriction in cancer growth control and life-span extension". Cancer Treatment Reviews. 38 (6): 726–36. doi:10.1016/j.ctrv.2012.01.004. PMID 22342103.
^ Gomez J, Caro P, Sanchez I, Naudi A, Jove M, Portero-Otin M, Lopez-Torres M, Pamplona R, Barja G (June 2009). "Effect of methionine dietary supplementation on mitochondrial oxygen radical generation and oxidative DNA damage in rat liver and heart". Journal of Bioenergetics and Biomembranes. 41 (3): 309–21. doi:10.1007/s10863-009-9229-3. PMID 19633937.
^ Oz HS, Chen TS, Neuman M (March 2008). "Methionine deficiency and hepatic injury in a dietary steatohepatitis model". Digestive Diseases and Sciences. 53 (3): 767–76. doi:10.1007/s10620-007-9900-7. PMC 2271115. PMID 17710550.
^ Yu D, Yang SE, Miller BR, Wisinski JA, Sherman DS, Brinkman JA, Tomasiewicz JL, Cummings NE, Kimple ME, Cryns VL, Lamming DW (January 2018). "Short-term methionine deprivation improves metabolic health via sexually dimorphic, mTORC1-independent mechanisms". FASEB Journal. 32 (6): 3471–3482. doi:10.1096/fj.201701211R. PMC 5956241. PMID 29401631.
^ Wood JM, Decker H, Hartmann H, Chavan B, Rokos H, Spencer JD, et al. (July 2009). "Senile hair graying: H2O2-mediated oxidative stress affects human hair color by blunting methionine sulfoxide repair". FASEB Journal. 23 (7): 2065–75. doi:10.1096/fj.08-125435. hdl:10454/6241. PMID 19237503.
^ Refsum H, Ueland PM, Nygård O, Vollset SE (1998). "Homocysteine and cardiovascular disease". Annual Review of Medicine. 49 (1): 31–62. doi:10.1146/annurev.med.49.1.31. PMID 9509248.
^ Weaver IC, Champagne FA, Brown SE, Dymov S, Sharma S, Meaney MJ, Szyf M (November 2005). "Reversal of maternal programming of stress responses in adult offspring through methyl supplementation: altering epigenetic marking later in life". The Journal of Neuroscience. 25 (47): 11045–54. doi:10.1523/JNEUROSCI.3652-05.2005. PMID 16306417.
^ Grimshaw, Jane (July 25, 2011) Methionine for Dogs uses and Side Effects. critters360.com
^ "Rules and Regulations". Federal Register. 76 (49): 13501–13504. March 14, 2011.
^ Lewis DS, Cuda JP, Stevens BR (December 2011). "A novel biorational pesticide: efficacy of methionine against Heraclides (Papilio) cresphontes, a surrogate of the invasive Princeps (Papilio) demoleus (Lepidoptera: Papilionidae)". Journal of Economic Entomology. 104 (6): 1986–90. PMID 22299361.
External links
Rudra MN, Chowdhury LM (30 September 1950). "Methionine Content of Cereals and Legumes". Nature. 166 (568): 568. Bibcode:1950Natur.166..568R. doi:10.1038/166568a0.
Encoded (proteinogenic) amino acids | |||||||||||
|---|---|---|---|---|---|---|---|---|---|---|---|
| General topics |
|  | |||||||||
| By properties |
| ||||||||||
| |||||||||||


