Molecular dynamics

Example of a molecular dynamics simulation in a simple system: deposition of one copper (Cu) atom on a Cu Miller index (001) surface. Each circle illustrates the position of one atom. The atomic interactions used in current simulations are more complex than those of 2-dimensional hard spheres.

Molecular dynamics is often used to simulate biophysical systems. Depicted here is a 100 ps simulation of water.
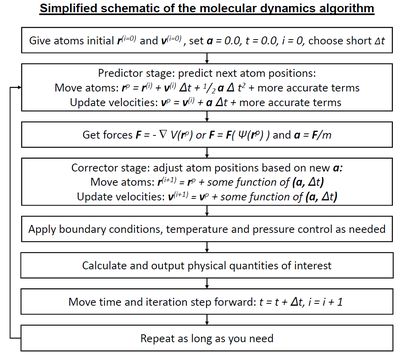
A simplified description of the standard molecular dynamics simulation algorithm, when a predictor-corrector-type integrator is used. The forces may come either from classical interatomic potentials (indicated as F=−∇V(r→){displaystyle F=-nabla V({vec {r}})}
 ) or quantum mechanical (indicated schematically as F=F(Ψ(r→)){displaystyle F=F(Psi ({vec {r}}))}
) or quantum mechanical (indicated schematically as F=F(Ψ(r→)){displaystyle F=F(Psi ({vec {r}}))} ) methods. Large differences exist between different integrators; some do not exactly have the same highest-order terms as indicated in the schematic, many use also higher-order time derivatives, and some use both the current and prior time step in variable-time step schemes.
) methods. Large differences exist between different integrators; some do not exactly have the same highest-order terms as indicated in the schematic, many use also higher-order time derivatives, and some use both the current and prior time step in variable-time step schemes.Molecular dynamics (MD) is a computer simulation method for studying the physical movements of atoms and molecules. The atoms and molecules are allowed to interact for a fixed period of time, giving a view of the dynamic evolution of the system. In the most common version, the trajectories of atoms and molecules are determined by numerically solving Newton's equations of motion for a system of interacting particles, where forces between the particles and their potential energies are often calculated using interatomic potentials or molecular mechanics force fields. The method was originally developed within the field of theoretical physics in the late 1950s[1][2][3] but is applied today mostly in chemical physics, materials science and the modelling of biomolecules.
Because molecular systems typically consist of a vast number of particles, it is impossible to determine the properties of such complex systems analytically; MD simulation circumvents this problem by using numerical methods. However, long MD simulations are mathematically ill-conditioned, generating cumulative errors in numerical integration that can be minimized with proper selection of algorithms and parameters, but not eliminated entirely.
For systems which obey the ergodic hypothesis, the evolution of one molecular dynamics simulation may be used to determine macroscopic thermodynamic properties of the system: the time averages of an ergodic system correspond to microcanonical ensemble averages. MD has also been termed "statistical mechanics by numbers" and "Laplace's vision of Newtonian mechanics" of predicting the future by animating nature's forces[4][5] and allowing insight into molecular motion on an atomic scale.
Contents
1 History
2 Areas of application and limits
3 Design constraints
3.1 Microcanonical ensemble (NVE)
3.2 Canonical ensemble (NVT)
3.3 Isothermal–isobaric (NPT) ensemble
3.4 Generalized ensembles
4 Potentials in MD simulations
4.1 Empirical potentials
4.2 Pair potentials versus many-body potentials
4.3 Semi-empirical potentials
4.4 Polarizable potentials
4.5 Potentials in ab initio methods
4.6 Hybrid QM/MM
4.7 Coarse-graining and reduced representations
5 Incorporating solvent effects
6 Long-range forces
7 Steered molecular dynamics (SMD)
8 Examples of applications
9 Molecular dynamics algorithms
9.1 Integrators
9.2 Short-range interaction algorithms
9.3 Long-range interaction algorithms
9.4 Parallelization strategies
10 Specialized hardware for MD simulations
11 Graphics card as a hardware for MD simulations
12 See also
13 References
13.1 General references
14 External links
History
Following the earlier successes of Monte Carlo simulations, the method was first developed by Fermi, Pasta, Ulam and Tsingou[1] in the mid 50s. In 1957, Alder and Wainwright used an IBM 704 computer to simulate perfectly elastic collisions between hard spheres.[2] In 1960, Gibson et al., simulated radiation damage of solid copper by using a Born–Mayer type of repulsive interaction along with a cohesive surface force.[6] In 1964, Rahman published landmark simulations of liquid argon that used a Lennard-Jones potential. Calculations of system properties, such as the coefficient of self-diffusion, compared well with experimental data.[3]
Even before it became possible to simulate molecular dynamics with computers, some undertook the hard work of trying it with physical models such as macroscopic spheres. The idea was to arrange them to replicate the properties of a liquid. J.D. Bernal said, in 1962: "... I took a number of rubber balls and stuck them together with rods of a selection of different lengths ranging from 2.75 to 4 inches. I tried to do this in the first place as casually as possible, working in my own office, being interrupted every five minutes or so and not remembering what I had done before the interruption."[7]
Areas of application and limits
Beginning in theoretical physics, the method of MD gained popularity in materials science and since the 1970s also in biochemistry and biophysics. MD is frequently used to refine 3-dimensional structures of proteins and other macromolecules based on experimental constraints from X-ray crystallography or NMR spectroscopy. In physics, MD is used to examine the dynamics of atomic-level phenomena that cannot be observed directly, such as thin film growth and ion-subplantation, and also to examine the physical properties of nanotechnological devices that have not or cannot yet be created. In biophysics and structural biology, the method is frequently applied to study the motions of macromolecules such as proteins and nucleic acids, which can be useful for interpreting the results of certain biophysical experiments and for modeling interactions with other molecules, as in ligand docking. In principle MD can be used for ab initio prediction of protein structure by simulating folding of the polypeptide chain from random coil.
The results of MD simulations can be tested through comparison to experiments that measure molecular dynamics, of which a popular method is NMR spectroscopy. MD-derived structure predictions can be tested through community-wide experiments in Critical Assessment of protein Structure Prediction (CASP), although the method has historically had limited success in this area. Michael Levitt, who shared the Nobel Prize partly for the application of MD to proteins, wrote in 1999 that CASP participants usually did not use the method due to "... a central embarrassment of molecular mechanics, namely that energy minimization or molecular dynamics generally leads to a model that is less like the experimental structure."[8] Improvements in computational resources permitting more and longer MD trajectories, combined with modern improvements in the quality of force field parameters, have yielded some improvements in both structure prediction and homology model refinement, without reaching the point of practical utility in these areas; many identify force field parameters as a key area for further development.[9][10][11]
Limits of the method are related to the parameter sets used, and to the underlying molecular mechanics force fields. One run of an MD simulation optimizes the potential energy, rather than the free energy of the protein[dubious ], meaning that all entropic contributions to thermodynamic stability of protein structure are neglected, including the conformational entropy of the polypeptide chain (the main factor that destabilizes protein structure) and hydrophobic effects (the main driving forces of protein folding).[12] Another important factor are intramolecular hydrogen bonds,[13] which are not explicitly included in modern force fields, but described as Coulomb interactions of atomic point charges. This is a crude approximation because hydrogen bonds have a partially quantum mechanical and chemical nature. Furthermore, electrostatic interactions are usually calculated using the dielectric constant of vacuum, although the surrounding aqueous solution has a much higher dielectric constant. Using the macroscopic dielectric constant at short interatomic distances is questionable. Finally, van der Waals interactions in MD are usually described by Lennard-Jones potentials based on the Fritz London theory that is only applicable in vacuum. However, all types of van der Waals forces are ultimately of electrostatic origin and therefore depend on dielectric properties of the environment.[14] The direct measurement of attraction forces between different materials (as Hamaker constant) shows that "the interaction between hydrocarbons across water is about 10% of that across vacuum".[14] The environment-dependence of van der Waals forces is neglected in standard simulations, but can be included by developing polarizable force fields.
Design constraints
Design of a molecular dynamics simulation should account for the available computational power. Simulation size (n = number of particles), timestep, and total time duration must be selected so that the calculation can finish within a reasonable time period. However, the simulations should be long enough to be relevant to the time scales of the natural processes being studied. To make statistically valid conclusions from the simulations, the time span simulated should match the kinetics of the natural process. Otherwise, it is analogous to making conclusions about how a human walks when only looking at less than one footstep. Most scientific publications about the dynamics of proteins and DNA [15][16] use data from simulations spanning nanoseconds (10−9 s) to microseconds (10−6 s). To obtain these simulations, several CPU-days to CPU-years are needed. Parallel algorithms allow the load to be distributed among CPUs; an example is the spatial or force decomposition algorithm.[17]
During a classical MD simulation, the most CPU intensive task is the evaluation of the potential as a function of the particles' internal coordinates. Within that energy evaluation, the most expensive one is the non-bonded or non-covalent part. In Big O notation, common molecular dynamics simulations scale by O(n2){displaystyle O(n^{2})}


Another factor that impacts total CPU time needed by a simulation is the size of the integration timestep. This is the time length between evaluations of the potential. The timestep must be chosen small enough to avoid discretization errors (i.e., smaller than the period related to fastest vibrational frequency in the system). Typical timesteps for classical MD are in the order of 1 femtosecond (10−15 s). This value may be extended by using algorithms such as the SHAKE constraint algorithm, which fix the vibrations of the fastest atoms (e.g., hydrogens) into place. Multiple time scale methods have also been developed, which allow extended times between updates of slower long-range forces.[18][19][20]
For simulating molecules in a solvent, a choice should be made between explicit and implicit solvent. Explicit solvent particles (such as the TIP3P, SPC/E and SPC-f water models) must be calculated expensively by the force field, while implicit solvents use a mean-field approach. Using an explicit solvent is computationally expensive, requiring inclusion of roughly ten times more particles in the simulation. But the granularity and viscosity of explicit solvent is essential to reproduce certain properties of the solute molecules. This is especially important to reproduce chemical kinetics.
In all kinds of molecular dynamics simulations, the simulation box size must be large enough to avoid boundary condition artifacts. Boundary conditions are often treated by choosing fixed values at the edges (which may cause artifacts), or by employing periodic boundary conditions in which one side of the simulation loops back to the opposite side, mimicking a bulk phase (which may cause artifacts too).
Microcanonical ensemble (NVE)
In the microcanonical ensemble, the system is isolated from changes in moles (N), volume (V), and energy (E). It corresponds to an adiabatic process with no heat exchange. A microcanonical molecular dynamics trajectory may be seen as an exchange of potential and kinetic energy, with total energy being conserved. For a system of N particles with coordinates X{displaystyle X}

- F(X)=−∇U(X)=MV˙(t){displaystyle F(X)=-nabla U(X)=M{dot {V}}(t)}
- V(t)=X˙(t).{displaystyle V(t)={dot {X}}(t).}


The potential energy function U(X){displaystyle U(X)}

For every time step, each particle's position X{displaystyle X}
One frequent source of confusion is the meaning of temperature in MD. Commonly we have experience with macroscopic temperatures, which involve a huge number of particles. But temperature is a statistical quantity. If there is a large enough number of atoms, statistical temperature can be estimated from the instantaneous temperature, which is found by equating the kinetic energy of the system to nkBT/2 where n is the number of degrees of freedom of the system.
A temperature-related phenomenon arises due to the small number of atoms that are used in MD simulations. For example, consider simulating the growth of a copper film starting with a substrate containing 500 atoms and a deposition energy of 100 eV. In the real world, the 100 eV from the deposited atom would rapidly be transported through and shared among a large number of atoms (1010{displaystyle 10^{10}}
Canonical ensemble (NVT)
In the canonical ensemble, amount of substance (N), volume (V) and temperature (T) are conserved. It is also sometimes called constant temperature molecular dynamics (CTMD). In NVT, the energy of endothermic and exothermic processes is exchanged with a thermostat.
A variety of thermostat algorithms are available to add and remove energy from the boundaries of a MD simulation in a more or less realistic way, approximating the canonical ensemble. Popular methods to control temperature include velocity rescaling, the Nosé–Hoover thermostat, Nosé–Hoover chains, the Berendsen thermostat, the Andersen thermostat and Langevin dynamics. The Berendsen thermostat might introduce the flying ice cube effect, which leads to unphysical translations and rotations of the simulated system.
It is not trivial to obtain a canonical ensemble distribution of conformations and velocities using these algorithms. How this depends on system size, thermostat choice, thermostat parameters, time step and integrator is the subject of many articles in the field.
Isothermal–isobaric (NPT) ensemble
In the isothermal–isobaric ensemble, amount of substance (N), pressure (P) and temperature (T) are conserved. In addition to a thermostat, a barostat is needed. It corresponds most closely to laboratory conditions with a flask open to ambient temperature and pressure.
In the simulation of biological membranes, isotropic pressure control is not appropriate. For lipid bilayers, pressure control occurs under constant membrane area (NPAT) or constant surface tension "gamma" (NPγT).
Generalized ensembles
The replica exchange method is a generalized ensemble. It was originally created to deal with the slow dynamics of disordered spin systems. It is also called parallel tempering. The replica exchange MD (REMD) formulation[21] tries to overcome the multiple-minima problem by exchanging the temperature of non-interacting replicas of the system running at several temperatures.
Potentials in MD simulations
A molecular dynamics simulation requires the definition of a potential function, or a description of the terms by which the particles in the simulation will interact. In chemistry and biology this is usually referred to as a force field and in materials physics as an interatomic potential. Potentials may be defined at many levels of physical accuracy; those most commonly used in chemistry are based on molecular mechanics and embody a classical mechanics treatment of particle-particle interactions that can reproduce structural and conformational changes but usually cannot reproduce chemical reactions.
The reduction from a fully quantum description to a classical potential entails two main approximations. The first one is the Born–Oppenheimer approximation, which states that the dynamics of electrons are so fast that they can be considered to react instantaneously to the motion of their nuclei. As a consequence, they may be treated separately. The second one treats the nuclei, which are much heavier than electrons, as point particles that follow classical Newtonian dynamics. In classical molecular dynamics, the effect of the electrons is approximated as one potential energy surface, usually representing the ground state.
When finer levels of detail are needed, potentials based on quantum mechanics are used; some methods attempt to create hybrid classical/quantum potentials where the bulk of the system is treated classically but a small region is treated as a quantum system, usually undergoing a chemical transformation.
Empirical potentials
Empirical potentials used in chemistry are frequently called force fields, while those used in materials physics are called interatomic potentials.
Most force fields in chemistry are empirical and consist of a summation of bonded forces associated with chemical bonds, bond angles, and bond dihedrals, and non-bonded forces associated with van der Waals forces and electrostatic charge. Empirical potentials represent quantum-mechanical effects in a limited way through ad-hoc functional approximations. These potentials contain free parameters such as atomic charge, van der Waals parameters reflecting estimates of atomic radius, and equilibrium bond length, angle, and dihedral; these are obtained by fitting against detailed electronic calculations (quantum chemical simulations) or experimental physical properties such as elastic constants, lattice parameters and spectroscopic measurements.
Because of the non-local nature of non-bonded interactions, they involve at least weak interactions between all particles in the system. Its calculation is normally the bottleneck in the speed of MD simulations. To lower the computational cost, force fields employ numerical approximations such as shifted cutoff radii, reaction field algorithms, particle mesh Ewald summation, or the newer particle–particle-particle–mesh (P3M).
Chemistry force fields commonly employ preset bonding arrangements (an exception being ab initio dynamics), and thus are unable to model the process of chemical bond breaking and reactions explicitly. On the other hand, many of the potentials used in physics, such as those based on the bond order formalism can describe several different coordinations of a system and bond breaking.[22][23] Examples of such potentials include the Brenner potential[24] for hydrocarbons and its
further developments for the C-Si-H[25] and C-O-H[26] systems. The
ReaxFF potential[27] can be considered a fully reactive hybrid between bond order potentials and chemistry force fields.
Pair potentials versus many-body potentials
The potential functions representing the non-bonded energy are formulated as a sum over interactions between the particles of the system. The simplest choice, employed in many popular force fields, is the "pair potential", in which the total potential energy can be calculated from the sum of energy contributions between pairs of atoms. An example of such a pair potential is the non-bonded Lennard–Jones potential (also termed the 6–12 potential), used for calculating van der Waals forces.
- U(r)=4ε[(σr)12−(σr)6]{displaystyle U(r)=4varepsilon left[left({frac {sigma }{r}}right)^{12}-left({frac {sigma }{r}}right)^{6}right]}
![<br />
U(r) = 4varepsilon left[ left(frac{sigma}{r}right)^{12} - left(frac{sigma}{r}right)^{6} right]<br />](https://wikimedia.org/api/rest_v1/media/math/render/svg/374024e23ac5eb77e91b68ad9ba86ad3bbf5f113)
Another example is the Born (ionic) model of the ionic lattice. The first term in the next equation is Coulomb's law for a pair of ions, the second term is the short-range repulsion explained by Pauli's exclusion principle and the final term is the dispersion interaction term. Usually, a simulation only includes the dipolar term, although sometimes the quadrupolar term is also included.[28][29] When nl = 6, this potential is also called the Coulomb–Buckingham potential.
- Uij(rij)=zizj4πϵ01rij+Alexp−rijpl+Clrij−nl+⋯{displaystyle U_{ij}(r_{ij})={frac {z_{i}z_{j}}{4pi epsilon _{0}}}{frac {1}{r_{ij}}}+A_{l}exp {frac {-r_{ij}}{p_{l}}}+C_{l}r_{ij}^{-n_{l}}+cdots }

In many-body potentials, the potential energy includes the effects of three or more particles interacting with each other.[30] In simulations with pairwise potentials, global interactions in the system also exist, but they occur only through pairwise terms. In many-body potentials, the potential energy cannot be found by a sum over pairs of atoms, as these interactions are calculated explicitly as a combination of higher-order terms. In the statistical view, the dependency between the variables cannot in general be expressed using only pairwise products of the degrees of freedom. For example, the Tersoff potential,[31] which was originally used to simulate carbon, silicon, and germanium, and has since been used for a wide range of other materials, involves a sum over groups of three atoms, with the angles between the atoms being an important factor in the potential. Other examples are the embedded-atom method (EAM),[32] the EDIP,[30] and the Tight-Binding Second Moment Approximation (TBSMA) potentials,[33] where the electron density of states in the region of an atom is calculated from a sum of contributions from surrounding atoms, and the potential energy contribution is then a function of this sum.
Semi-empirical potentials
Semi-empirical potentials make use of the matrix representation from quantum mechanics. However, the values of the matrix elements are found through empirical formulae that estimate the degree of overlap of specific atomic orbitals. The matrix is then diagonalized to determine the occupancy of the different atomic orbitals, and empirical formulae are used once again to determine the energy contributions of the orbitals.
There are a wide variety of semi-empirical potentials, termed tight-binding potentials, which vary according to the atoms being modeled.
Polarizable potentials
Most classical force fields implicitly include the effect of polarizability, e.g., by scaling up the partial charges obtained from quantum chemical calculations. These partial charges are stationary with respect to the mass of the atom. But molecular dynamics simulations can explicitly model polarizability with the introduction of induced dipoles through different methods, such as Drude particles or fluctuating charges. This allows for a dynamic redistribution of charge between atoms which responds to the local chemical environment.
For many years, polarizable MD simulations have been touted as the next generation. For homogenous liquids such as water, increased accuracy has been achieved through the inclusion of polarizability.[34][35][36] Some promising results have also been achieved for proteins.[37][38] However, it is still uncertain how to best approximate polarizability in a simulation.[citation needed]
Potentials in ab initio methods
In classical molecular dynamics, one potential energy surface (usually the ground state) is represented in the force field. This is a consequence of the Born–Oppenheimer approximation. In excited states, chemical reactions or when a more accurate representation is needed, electronic behavior can be obtained from first principles by using a quantum mechanical method, such as density functional theory. This is named Ab Initio Molecular Dynamics (AIMD). Due to the cost of treating the electronic degrees of freedom, the computational cost of these simulations is far higher than classical molecular dynamics. This implies that AIMD is limited to smaller systems and shorter times.
Ab initio quantum mechanical and chemical methods may be used to calculate the potential energy of a system on the fly, as needed for conformations in a trajectory. This calculation is usually made in the close neighborhood of the reaction coordinate. Although various approximations may be used, these are based on theoretical considerations, not on empirical fitting. Ab initio calculations produce a vast amount of information that is not available from empirical methods, such as density of electronic states or other electronic properties. A significant advantage of using ab initio methods is the ability to study reactions that involve breaking or formation of covalent bonds, which correspond to multiple electronic states.
Hybrid QM/MM
QM (quantum-mechanical) methods are very powerful. However, they are computationally expensive, while the MM (classical or molecular mechanics) methods are fast but suffer from several limits (require extensive parameterization; energy estimates obtained are not very accurate; cannot be used to simulate reactions where covalent bonds are broken/formed; and are limited in their abilities for providing accurate details regarding the chemical environment). A new class of method has emerged that combines the good points of QM (accuracy) and MM (speed) calculations. These methods are termed mixed or hybrid quantum-mechanical and molecular mechanics methods (hybrid QM/MM).[39]
The most important advantage of hybrid QM/MM method is the speed. The cost of doing classical molecular dynamics (MM) in the most straightforward case scales O(n2), where n is the number of atoms in the system. This is mainly due to electrostatic interactions term (every particle interacts with every other particle). However, use of cutoff radius, periodic pair-list updates and more recently the variations of the particle-mesh Ewald's (PME) method has reduced this to between O(n) to O(n2). In other words, if a system with twice as many atoms is simulated then it would take between two and four times as much computing power. On the other hand, the simplest ab initio calculations typically scale O(n3) or worse (restricted Hartree–Fock calculations have been suggested to scale ~O(n2.7)). To overcome the limit, a small part of the system is treated quantum-mechanically (typically active-site of an enzyme) and the remaining system is treated classically.
In more sophisticated implementations, QM/MM methods exist to treat both light nuclei susceptible to quantum effects (such as hydrogens) and electronic states. This allows generating hydrogen wave-functions (similar to electronic wave-functions). This methodology has been useful in investigating phenomena such as hydrogen tunneling. One example where QM/MM methods have provided new discoveries is the calculation of hydride transfer in the enzyme liver alcohol dehydrogenase. In this case, quantum tunneling is important for the hydrogen, as it determines the reaction rate.[40]
Coarse-graining and reduced representations
At the other end of the detail scale are coarse-grained and lattice models. Instead of explicitly representing every atom of the system, one uses "pseudo-atoms" to represent groups of atoms. MD simulations on very large systems may require such large computer resources that they cannot easily be studied by traditional all-atom methods. Similarly, simulations of processes on long timescales (beyond about 1 microsecond) are prohibitively expensive, because they require so many time steps. In these cases, one can sometimes tackle the problem by using reduced representations, which are also called coarse-grained models.[41]
Examples for coarse graining (CG) methods are discontinuous molecular dynamics (CG-DMD)[42][43] and Go-models.[44] Coarse-graining is done sometimes taking larger pseudo-atoms. Such united atom approximations have been used in MD simulations of biological membranes. Implementation of such approach on systems where electrical properties are of interest can be challenging owing to the difficulty of using a proper charge distribution on the pseudo-atoms.[45] The aliphatic tails of lipids are represented by a few pseudo-atoms by gathering 2 to 4 methylene groups into each pseudo-atom.
The parameterization of these very coarse-grained models must be done empirically, by matching the behavior of the model to appropriate experimental data or all-atom simulations. Ideally, these parameters should account for both enthalpic and entropic contributions to free energy in an implicit way. When coarse-graining is done at higher levels, the accuracy of the dynamic description may be less reliable. But very coarse-grained models have been used successfully to examine a wide range of questions in structural biology, liquid crystal organization, and polymer glasses.
Examples of applications of coarse-graining:
protein folding and protein structure prediction studies are often carried out using one, or a few, pseudo-atoms per amino acid;[41]
liquid crystal phase transitions have been examined in confined geometries and/or during flow using the Gay-Berne potential, which describes anisotropic species;
Polymer glasses during deformation have been studied using simple harmonic or FENE springs to connect spheres described by the Lennard-Jones potential;
DNA supercoiling has been investigated using 1–3 pseudo-atoms per basepair, and at even lower resolution;- Packaging of double-helical DNA into bacteriophage has been investigated with models where one pseudo-atom represents one turn (about 10 basepairs) of the double helix;
- RNA structure in the ribosome and other large systems has been modeled with one pseudo-atom per nucleotide.
- Virtual cell simulation to study the interaction of cells and various substrates.[46]
The simplest form of coarse-graining is the united atom (sometimes called extended atom) and was used in most early MD simulations of proteins, lipids, and nucleic acids. For example, instead of treating all four atoms of a CH3 methyl group explicitly (or all three atoms of CH2 methylene group), one represents the whole group with one pseudo-atom. It must, of course, be properly parameterized so that its van der Waals interactions with other groups have the proper distance-dependence. Similar considerations apply to the bonds, angles, and torsions in which the pseudo-atom participates. In this kind of united atom representation, one typically eliminates all explicit hydrogen atoms except those that have the capability to participate in hydrogen bonds (polar hydrogens). An example of this is the CHARMM 19 force-field.
The polar hydrogens are usually retained in the model, because proper treatment of hydrogen bonds requires a reasonably accurate description of the directionality and the electrostatic interactions between the donor and acceptor groups. A hydroxyl group, for example, can be both a hydrogen bond donor, and a hydrogen bond acceptor, and it would be impossible to treat this with one OH pseudo-atom. About half the atoms in a protein or nucleic acid are non-polar hydrogens, so the use of united atoms can provide a substantial savings in computer time.
Incorporating solvent effects
In many simulations of a solute-solvent system the main focus is on the behavior of the solute with little interest of the solvent behavior particularly in those solvent molecules residing in regions far from the solute molecule.[47] Solvents may influence the dynamic behavior of solutes via random collisions and by imposing a frictional drag on the motion of the solute through the solvent. The use of non-rectangular periodic boundary conditions, stochastic boundaries and solvent shells can all help reduce the number of solvent molecules required and enable a larger proportion of the computing time to be spent instead on simulating the solute. It is also possible to incorporate the effects of a solvent without needing any explicit solvent molecules present. One example of this approach is to use a potential mean force (PMF) which describes how the free energy changes as a particular coordinate is varied. The free energy change described by PMF contains the averaged effects of the solvent.
Long-range forces
A long range interaction is an interaction in which the spatial interaction falls off no faster than r−d{displaystyle r^{-d}}

Steered molecular dynamics (SMD)
Steered molecular dynamics (SMD) simulations, or force probe simulations, apply forces to a protein in order to manipulate its structure by pulling it along desired degrees of freedom. These experiments can be used to reveal structural changes in a protein at the atomic level. SMD is often used to simulate events such as mechanical unfolding or stretching.[49]
There are two typical protocols of SMD: one in which pulling velocity is held constant, and one in which applied force is constant. Typically, part of the studied system (e.g., an atom in a protein) is restrained by a harmonic potential. Forces are then applied to specific atoms at either a constant velocity or a constant force. Umbrella sampling is used to move the system along the desired reaction coordinate by varying, for example, the forces, distances, and angles manipulated in the simulation. Through umbrella sampling, all of the system's configurations—both high-energy and low-energy—are adequately sampled. Then, each configuration's change in free energy can be calculated as the potential of mean force.[50] A popular method of computing PMF is through the weighted histogram analysis method (WHAM), which analyzes a series of umbrella sampling simulations.[51][52]
Examples of applications

Molecular dynamics simulation of a synthetic molecular motor composed of three molecules in a nanopore (outer diameter 6.7 nm) at 250 K.
Molecular dynamics is used in many fields of science.
- First MD simulation of a simplified biological folding process was published in 1975. Its simulation published in Nature paved the way for the vast area of modern computational protein-folding.[53]
- First MD simulation of a biological process was published in 1976. Its simulation published in Nature paved the way for understanding protein motion as essential in function and not just accessory.[54]
- MD is the standard method to treat collision cascades in the heat spike regime, i.e., the effects that energetic neutron and ion irradiation have on solids and solid surfaces.[55][56]
- MD simulations were successfully applied to predict the molecular basis of the most common protein mutation N370S, causing Gaucher Disease.[57] In a follow-up publication it was shown that these blind predictions show a high correlation with experimental work on the same mutant, published independently at a later point.[58]
- MD simulations have been used to investigate the effect of surface charges on disjoining pressure of thin water films on metal surfaces.[59]
- MD simulations are used along with multislice image simulations to understand transmission electron microscope image features[60]
- MD calculations coupled with Hamiltonian-biasing algorithms have been used to probe the encapsulation thermodynamics of DNA double-strands onto hydrophobic[15] and hydrophilic[16] single-walled carbon nanotubes.
The following biophysical examples illustrate notable efforts to produce simulations of a systems of very large size (a complete virus) or very long simulation times (up to 1.112 milliseconds):
- MD simulation of the full satellite tobacco mosaic virus (STMV) (2006, Size: 1 million atoms, Simulation time: 50 ns, program: NAMD) This virus is a small, icosahedral plant virus that worsens the symptoms of infection by Tobacco Mosaic Virus (TMV). Molecular dynamics simulations were used to probe the mechanisms of viral assembly. The entire STMV particle consists of 60 identical copies of one protein that make up the viral capsid (coating), and a 1063 nucleotide single stranded RNA genome. One key finding is that the capsid is very unstable when there is no RNA inside. The simulation would take one 2006 desktop computer around 35 years to complete. It was thus done in many processors in parallel with continuous communication between them.[61]
- Folding simulations of the Villin Headpiece in all-atom detail (2006, Size: 20,000 atoms; Simulation time: 500 µs= 500,000 ns, Program: Folding@home) This simulation was run in 200,000 CPU's of participating personal computers around the world. These computers had the Folding@home program installed, a large-scale distributed computing effort coordinated by Vijay Pande at Stanford University. The kinetic properties of the Villin Headpiece protein were probed by using many independent, short trajectories run by CPU's without continuous real-time communication. One method employed was the Pfold value analysis, which measures the probability of folding before unfolding of a specific starting conformation. Pfold gives information about transition state structures and an ordering of conformations along the folding pathway. Each trajectory in a Pfold calculation can be relatively short, but many independent trajectories are needed.[62]
- Long continuous-trajectory simulations have been performed on Anton, a massively parallel supercomputer designed and built around custom application-specific integrated circuits (ASICs) and interconnects by D. E. Shaw Research. The longest published result of a simulation performed using Anton is a 1.112-millisecond simulation of NTL9 at 355 K; a second, independent 1.073-millisecond simulation of this configuration was also performed (and many other simulations of over 250 µs continuous chemical time).[63] In How Fast-Folding Proteins Fold, researchers Kresten Lindorff-Larsen, Stefano Piana, Ron O. Dror, and David E. Shaw discuss "the results of atomic-level molecular dynamics simulations, over periods ranging between 100 μs and 1 ms, that reveal a set of common principles underlying the folding of 12 structurally diverse proteins." Examination of these diverse long trajectories, enabled by specialized, custom hardware, allow them to conclude that "In most cases, folding follows a single dominant route in which elements of the native structure appear in an order highly correlated with their propensity to form in the unfolded state."[63] In a separate study, Anton was used to conduct a 1.013-millisecond simulation of the native-state dynamics of bovine pancreatic trypsin inhibitor (BPTI) at 300 K.[64]
- These molecular simulations have been used to understand the material removal mechanisms, effects of tool geometry, temperature, and process parameters such as cutting speed and cutting forces.[65] It was also used to investigate the mechanisms behind the exfoliation of few layers of graphene[66][67] and carbon nanoscrolls.
Molecular dynamics algorithms
- Screened Coulomb Potentials Implicit Solvent Model
Integrators
- Symplectic integrator
- Verlet-Stoermer integration
- Runge–Kutta integration
- Beeman's algorithm
Constraint algorithms (for constrained systems)
Short-range interaction algorithms
- Cell lists
- Verlet list
- Bonded interactions
Long-range interaction algorithms
- Ewald summation
- Particle mesh Ewald summation (PME)
- Particle–particle-particle–mesh (P3M)
- Shifted force method
Parallelization strategies
Domain decomposition method (Distribution of system data for parallel computing)
Specialized hardware for MD simulations
Anton – A specialized, massively parallel supercomputer designed to execute MD simulations
MDGRAPE – A special purpose system built for molecular dynamics simulations, especially protein structure prediction
Graphics card as a hardware for MD simulations
- Molecular modeling on GPU
See also
- Molecular modeling
- Computational chemistry
- Force field (chemistry)
- Comparison of force field implementations
- Monte Carlo method
- Molecular design software
- Molecular mechanics
- Multiscale Green's function
- Car–Parrinello method
- Comparison of software for molecular mechanics modeling
- Quantum chemistry
- Discrete element method
- Comparison of nucleic acid simulation software
- Molecule editor
References
^ ab Fermi E., Pasta J., Ulam S., Los Alamos report LA-1940 (1955).
^ ab Alder, B. J.; Wainwright, T. E. (1959). "Studies in Molecular Dynamics. I. General Method". J. Chem. Phys. 31 (2): 459. Bibcode:1959JChPh..31..459A. doi:10.1063/1.1730376..mw-parser-output cite.citation{font-style:inherit}.mw-parser-output q{quotes:"""""""'""'"}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
^ ab Rahman, A. (19 October 1964). "Correlations in the Motion of Atoms in Liquid Argon". Physical Review. 136 (2A): A405–A411. Bibcode:1964PhRv..136..405R. doi:10.1103/PhysRev.136.A405.
^ Schlick, T. (1996). "Pursuing Laplace's Vision on Modern Computers". In J. P. Mesirov, K. Schulten and D. W. Sumners. Mathematical Applications to Biomolecular Structure and Dynamics, IMA Volumes in Mathematics and Its Applications. 82. New York: Springer-Verlag. pp. 218–247. ISBN 978-0-387-94838-6.
^ de Laplace, P. S. (1820). Oeuveres Completes de Laplace, Theorie Analytique des Probabilites (in French). Paris, France: Gauthier-Villars.
^ Gibson, J B; Goland, A N; Milgram, M; Vineyard, G H (1960). "Dynamics of Radiation Damage". Phys. Rev. 120 (4): 1229–1253. Bibcode:1960PhRv..120.1229G. doi:10.1103/PhysRev.120.1229.
^ Bernal, J.D. (1964). "The Bakerian lecture, 1962: The structure of liquids". Proceedings of the Royal Society. 280 (1382): 299–322. Bibcode:1964RSPSA.280..299B. doi:10.1098/rspa.1964.0147.
^ Koehl, P.; Levitt, Michael (1999). "A brighter future for protein structure prediction". Nature Structural Biology. 6: 108–111.
^ Raval, A; Piana, S; Eastwood, MP; Dror, RO; Shaw, DE (August 2012). "Refinement of protein structure homology models via long, all-atom molecular dynamics simulations". Proteins. 80 (8): 2071–9. doi:10.1002/prot.24098. PMID 22513870.
^ Beauchamp, KA; Lin, YS; Das, R; Pande, VS (10 April 2012). "Are Protein Force Fields Getting Better? A Systematic Benchmark on 524 Diverse NMR Measurements". Journal of Chemical Theory and Computation. 8 (4): 1409–1414. doi:10.1021/ct2007814. PMC 3383641. PMID 22754404.
^ Piana, S; Klepeis, JL; Shaw, DE (February 2014). "Assessing the accuracy of physical models used in protein-folding simulations: quantitative evidence from long molecular dynamics simulations". Current Opinion in Structural Biology. 24: 98–105. doi:10.1016/j.sbi.2013.12.006. PMID 24463371.
^ Hydrophobic effects are mostly of entropic nature at room temperature.
^ Myers, J. K.; Pace, C. N. (1996). "Hydrogen bonding stabilizes globular proteins". Biophys. J. 71 (4): 2033–2039. Bibcode:1996BpJ....71.2033M. doi:10.1016/s0006-3495(96)79401-8. PMC 1233669. PMID 8889177.
^ ab Israelachvili, Jacob (1992). Intermolecular and surface forces. Academic Press, San Diego.
^ ab Cruz, F.J.A.L.; de Pablo, J.J.; Mota, J.P.B. (2014), "Endohedral confinement of a DNA dodecamer onto pristine carbon nanotubes and the stability of the canonical B form", J. Chem. Phys., 140: 225103, arXiv:1605.01317, Bibcode:2014JChPh.140v5103C, doi:10.1063/1.4881422
^ ab Cruz, F.J.A.L.; Mota, J.P.B. (2016), "Conformational Thermodynamics of DNA Strands in Hydrophilic Nanopores", J. Phys. Chem. C, 120: 20357–20367, doi:10.1021/acs.jpcc.6b06234
^ Plimpton, Steve. Molecular Dynamics - Parallel Algorithms. sandia.gov
^ Streett WB, Tildesley DJ, Saville G; Tildesley; Saville (1978). "Multiple time-step methods in molecular dynamics". Mol Phys. 35 (3): 639–648. Bibcode:1978MolPh..35..639S. doi:10.1080/00268977800100471.CS1 maint: Multiple names: authors list (link)
^ Tuckerman ME, Berne BJ, Martyna GJ; Berne; Martyna (1991). "Molecular dynamics algorithm for multiple time scales: systems with long range forces". J Chem Phys. 94 (10): 6811–6815. Bibcode:1991JChPh..94.6811T. doi:10.1063/1.460259.CS1 maint: Multiple names: authors list (link)
^ Tuckerman ME, Berne BJ, Martyna GJ; Berne; Martyna (1992). "Reversible multiple time scale molecular dynamics". J Chem Phys. 97 (3): 1990–2001. Bibcode:1992JChPh..97.1990T. doi:10.1063/1.463137.CS1 maint: Multiple names: authors list (link)
^ Sugita, Yuji; Yuko Okamoto (1999). "Replica-exchange molecular dynamics method for protein folding". Chem Phys Lett. 314 (1): 141–151. Bibcode:1999CPL...314..141S. doi:10.1016/S0009-2614(99)01123-9.
^ Sinnott, S. B.; Brenner, D. W. (2012). "Three decades of many-body potentials in materials research". MRS Bulletin. 37 (5): 469–473. doi:10.1557/mrs.2012.88.
^ Albe, K.; Nordlund, K.; Averback, R. S. (2002). "Modeling metal-semiconductor interaction: Analytical bond-order potential for platinum-carbon". Phys. Rev. B. 65 (19): 195124. Bibcode:2002PhRvB..65s5124A. doi:10.1103/physrevb.65.195124.
^ Brenner, D. W. (1990). "Empirical potential for hydrocarbons for use in simulating the chemical vapor deposition of diamond films". Phys. Rev. B. 42 (15): 9458–9471. Bibcode:1990PhRvB..42.9458B. doi:10.1103/PhysRevB.42.9458.
^ Beardmore, Keith; Smith, Roger (1996). "Empirical potentials for C-Si-H systems with application to C60 interactions with Si crystal surfaces". Philosophical Magazine A. 74 (6): 1439–1466. Bibcode:1996PMagA..74.1439B. doi:10.1080/01418619608240734.
^ Ni, Boris; Lee, Ki-Ho; Sinnott, Susan B (2004). "A reactive empirical bond order (rebo) potential for hydrocarbon oxygen interactions". Journal of Physics: Condensed Matter. 16 (41): 7261–7275. Bibcode:2004JPCM...16.7261N. doi:10.1088/0953-8984/16/41/008.
^ van Duin, A.; Siddharth Dasgupta, François Lorant and William A. Goddard III; Lorant, Francois; Goddard, William A. (2001). "ReaxFF: A Reactive Force Field for Hydrocarbons". J. Phys. Chem. A. 105 (41): 9398. Bibcode:2001JPCA..105.9396V. CiteSeerX 10.1.1.507.6992. doi:10.1021/jp004368u.
^ Cruz, F.J.A.L.; Canongia Lopes, J.N.; Calado, J.C.G.; Minas da Piedade, M.E. (2005). "A Molecular Dynamics Study of the Thermodynamic Properties of Calcium Apatites. 1. Hexagonal Phases". J. Phys. Chem. B. 109 (51): 24473–24479. doi:10.1021/jp054304p. PMID 16375450.
^ Cruz, F.J.A.L.; Canongia Lopes, J.N.; Calado, J.C.G. (2006). "Molecular dynamics simulations of molten calcium hydroxyapatite". Fluid Phase Eq. 241 (1–2): 51–58. doi:10.1016/j.fluid.2005.12.021.
^ ab Justo, J. F.; Bazant, M. Z.; Kaxiras, E.; Bulatov, V. V.; Yip, S. (1998). "Interatomic potential for silicon defects and disordered phases". Phys. Rev. B. 58 (5): 2539–2550. arXiv:cond-mat/9712058. Bibcode:1998PhRvB..58.2539J. doi:10.1103/PhysRevB.58.2539.
^ Tersoff, J. (1989). "Modeling solid-state chemistry: Interatomic potentials for multicomponent systems". Phys. Rev. B. 39 (8): 5566–5568. Bibcode:1989PhRvB..39.5566T. doi:10.1103/PhysRevB.39.5566.
^ Daw, M. S.; S. M. Foiles and M. I. Baskes (1993). "The embedded-atom method: a review of theory and applications". Mat. Sci. And Engr. Rep. (Submitted manuscript). 9 (7–8): 251–310. doi:10.1016/0920-2307(93)90001-U.
^ Cleri, F.; V. Rosato (1993). "Tight-binding potentials for transition metals and alloys". Phys. Rev. B. 48 (1): 22–33. Bibcode:1993PhRvB..48...22C. doi:10.1103/PhysRevB.48.22.
^ Lamoureux G, Harder E, Vorobyov IV, Roux B, MacKerell AD; Harder; Vorobyov; Roux; MacKerell (2006). "A polarizable model of water for molecular dynamics simulations of biomolecules". Chem Phys Lett. 418 (1): 245–249. Bibcode:2006CPL...418..245L. doi:10.1016/j.cplett.2005.10.135.CS1 maint: Multiple names: authors list (link)
^ Sokhan VP, Jones AP, Cipcigan FS, Crain J, Martyna GJ (2015). "Signature properties of water: Their molecular electronic origins". Proceedings of the National Academy of Sciences. 112 (20): 6341–6346. Bibcode:2015PNAS..112.6341S. doi:10.1073/pnas.1418982112. PMC 4443379. PMID 25941394.
^ Cipcigan FS, Sokhan VP, Jones AP, Crain J, Martyna GJ (2015). "Hydrogen bonding and molecular orientation at the liquid–vapour interface of water". Physical Chemistry Chemical Physics. 17 (14): 8660–8669. Bibcode:2015PCCP...17.8660C. doi:10.1039/C4CP05506C. PMID 25715668.
^ Mahmoudi, Morteza; Lynch, Iseult; Ejtehadi, Mohammad Reza; Monopoli, Marco P.; Bombelli, Francesca Baldelli; Laurent, Sophie (2011). "Protein−Nanoparticle Interactions: Opportunities and Challenges". Chemical Reviews. 111 (9): 5610–37. doi:10.1021/cr100440g. PMID 21688848.
^ Patel, S.; MacKerell, Jr. AD; Brooks III, Charles L (2004). "CHARMM fluctuating charge force field for proteins: II protein/solvent properties from molecular dynamics simulations using a nonadditive electrostatic model". J Comput Chem. 25 (12): 1504–1514. doi:10.1002/jcc.20077. PMID 15224394.
^ The methodology for such methods was introduced by Warshel and coworkers. In the recent years have been pioneered by several groups including: Arieh Warshel (University of Southern California), Weitao Yang (Duke University), Sharon Hammes-Schiffer (The Pennsylvania State University), Donald Truhlar and Jiali Gao (University of Minnesota) and Kenneth Merz (University of Florida).
^ Billeter, SR; SP Webb; PK Agarwal; T Iordanov; S Hammes-Schiffer (2001). "Hydride Transfer in Liver Alcohol Dehydrogenase: Quantum Dynamics, Kinetic Isotope Effects, and Role of Enzyme Motion". J Am Chem Soc. 123 (45): 11262–11272. doi:10.1021/ja011384b. PMID 11697969.
^ ab Kmiecik, Sebastian; Gront, Dominik; Kolinski, Michal; Wieteska, Lukasz; Dawid, Aleksandra Elzbieta; Kolinski, Andrzej (2016-06-22). "Coarse-Grained Protein Models and Their Applications". Chemical Reviews. 116 (14): 7898–936. doi:10.1021/acs.chemrev.6b00163. ISSN 0009-2665. PMID 27333362.
^ Smith, A; CK Hall (2001). "Alpha-Helix Formation: Discontinuous Molecular Dynamics on an Intermediate-Resolution Protein Model". Proteins. 44 (3): 344–360. doi:10.1002/prot.1100. PMID 11455608.
^ Ding, F; JM Borreguero; SV Buldyrey; HE Stanley; NV Dokholyan (2003). "Mechanism for the alpha-helix to beta-hairpin transition". J Am Chem Soc. 53 (2): 220–228. doi:10.1002/prot.10468. PMID 14517973.
^ Paci, E; M Vendruscolo; M Karplus (2002). "Validity of Go Models: Comparison with a Solvent-Shielded Empirical Energy Decomposition". Biophys J. 83 (6): 3032–3038. Bibcode:2002BpJ....83.3032P. doi:10.1016/S0006-3495(02)75308-3. PMC 1302383. PMID 12496075.
^ Chakrabarty, A; T Cagin (2010). "Coarse grain modeling of polyimide copolymers". Polymer. 51 (12): 2786–2794. doi:10.1016/j.polymer.2010.03.060.
^ Heydari, Tiam; Heidari, Maziar; Mashinchian, Omid; Wojcik, Michal; Xu, Ke; Dalby, Matthew John; Mahmoudi, Morteza; Ejtehadi, Mohammad Reza (2017). "Development of a Virtual Cell Model to Predict Cell Response to Substrate Topography". ACS Nano. 11 (9): 9084–9092. doi:10.1021/acsnano.7b03732. PMID 28742318.
^ Leach, Dr Andrew (30 January 2001). Molecular Modelling: Principles and Applications (2nd ed.). Harlow: Prentice Hall. ISBN 9780582382107. ASIN 0582382106.
^ Allen, Michael P.; Tildesley, Dominic J. (2017-08-22). Computer Simulation of Liquids (2nd ed.). Oxford University Press. p. 216. ISBN 9780198803201. ASIN 0198803206.
^ Nienhaus, Gerd Ulrich (2005). Protein-ligand interactions: methods and applications. pp. 54–56. ISBN 978-1-61737-525-5.
^ Leszczyński, Jerzy (2005). Computational chemistry: reviews of current trends, Volume 9. pp. 54–56. ISBN 978-981-256-742-0.
^ Kumar, Shankar; Rosenberg, John M.; Bouzida, Djamal; Swendsen, Robert H.; Kollman, Peter A. (30 September 1992). "The weighted histogram analysis method for free-energy calculations on biomolecules. I. The method". Journal of Computational Chemistry. 13 (8): 1011–1021. doi:10.1002/jcc.540130812.
^ Bartels, Christian (1 December 2000). "Analyzing biased Monte Carlo and molecular dynamics simulations". Chemical Physics Letters. 331 (5–6): 446–454. Bibcode:2000CPL...331..446B. doi:10.1016/S0009-2614(00)01215-X.
^ Levitt, M; A Warshel (1975). "Computer Simulations of Protein Folding". Nature. 253 (5494): 694–8. Bibcode:1975Natur.253..694L. doi:10.1038/253694a0. PMID 1167625.
^ Warshel, A (1976). "Bicycle-pedal Model for the First Step in the Vision Process". Nature. 260 (5553): 679–683. Bibcode:1976Natur.260..694B. doi:10.1038/260679a0. PMID 1264239.
^ Averback, R. S.; Diaz de la Rubia, T. (1998). "Displacement damage in irradiated metals and semiconductors". In H. Ehrenfest and F. Spaepen. Solid State Physics. 51. New York: Academic Press. pp. 281–402.
^ Smith, R., ed. (1997). Atomic & ion collisions in solids and at surfaces: theory, simulation and applications. Cambridge, UK: Cambridge University Press.
^ Offman, MN; M Krol; I Silman; JL Sussman; AH Futerman (2010). "Molecular basis of reduced glucosylceramidase activity in the most common Gaucher disease mutant, N370S". J. Biol. Chem. 285 (53): 42105–42114. doi:10.1074/jbc.M110.172098. PMC 3009936. PMID 20980259.
^ Offman, MN; M Krol; B Rost; I Silman; JL Sussman; AH Futerman (2011). "Comparison of a molecular dynamics model with the X-ray structure of the N370S acid-beta-glucosidase mutant that causes Gaucher disease". Protein Eng. Des. Sel. 24 (10): 773–775. doi:10.1093/protein/gzr032. PMID 21724649.
^ Hu, Han; Sun, Ying (2013). "Molecular dynamics simulations of disjoining pressure effect in ultra-thin water film on a metal surface". Appl. Phys. Lett. 14 (26): 263110. Bibcode:2013ApPhL.103z3110H. doi:10.1063/1.4858469.
^ Welch, D. A.; Mehdi, B. L.; Hatchell, H. J.; Faller, R.; Evans, J. E.; Browning, N. D. (2015). "Using molecular dynamics to quantify the electrical double layer and examine the potential for its direct observation in the in-situ TEM". Advanced Structural and Chemical Imaging. 1. doi:10.1186/s40679-014-0002-2.
^ Freddolino P, Arkhipov A, Larson SB, McPherson A, Schulten K. "Molecular dynamics simulation of the Satellite Tobacco Mosaic Virus (STMV)". Theoretical and Computational Biophysics Group. University of Illinois at Urbana Champaign.
^ Jayachandran, Guha; Vishal, V.; Pande, Vijay S. (28 April 2006). "Using massively parallel simulation and Markovian models to study protein folding: Examining the dynamics of the villin headpiece". The Journal of Chemical Physics. 124 (16): 164902. Bibcode:2006JChPh.124p4902J. doi:10.1063/1.2186317. PMID 16674165.
^ ab Lindorff-Larsen, Kresten; Piana, Stefano; Dror, Ron O.; Shaw, David E. (2011). "How Fast-Folding Proteins Fold". Science. 334 (6055): 517–520. Bibcode:2011Sci...334..517L. CiteSeerX 10.1.1.1013.9290. doi:10.1126/science.1208351. PMID 22034434.
^ Shaw, David E.; Maragakis, Paul; Lindorff-Larsen, Kresten; Piana, Stefano; Dror, Ron O.; Eastwood, Michael P.; Bank, Joseph A.; Jumper, John M.; Salmon, John K.; et al. (2010). "Atomic-Level Characterization of the Structural Dynamics of Proteins". Science. 330 (6002): 341–346. Bibcode:2010Sci...330..341S. doi:10.1126/science.1187409. PMID 20947758.
^ Goel S, Luo; Reuben R L (2012). "Molecular dynamics simulation model for the quantitative assessment of tool wear during single point diamond turning of cubic silicon carbide". Comput. Mater. Sci. 51: 402–408. doi:10.1016/j.commatsci.2011.07.052.
^ Jayasena, Buddhika; Subbiah Sathyan (2011). "A novel mechanical cleavage method for synthesizing few-layer graphenes". Nanoscale Research Letters. 6 (1): 95. Bibcode:2011NRL.....6...95J. doi:10.1186/1556-276X-6-95. PMC 3212245. PMID 21711598.
^ Jayasena, B; Reddy C.D; Subbiah S (2013). "Separation, folding and shearing of graphene layers during wedge-based mechanical exfoliation". Nanotechnology. 24 (20): 205301. Bibcode:2013Nanot..24t5301J. doi:10.1088/0957-4484/24/20/205301. PMID 23598423.
General references
.mw-parser-output .refbegin{font-size:90%;margin-bottom:0.5em}.mw-parser-output .refbegin-hanging-indents>ul{list-style-type:none;margin-left:0}.mw-parser-output .refbegin-hanging-indents>ul>li,.mw-parser-output .refbegin-hanging-indents>dl>dd{margin-left:0;padding-left:3.2em;text-indent:-3.2em;list-style:none}.mw-parser-output .refbegin-100{font-size:100%}
- M. P. Allen, D. J. Tildesley (1989) Computer simulation of liquids. Oxford University Press.
ISBN 0-19-855645-4. - J. A. McCammon, S. C. Harvey (1987) Dynamics of Proteins and Nucleic Acids. Cambridge University Press.
ISBN 0-521-30750-3 (hardback). - D. C. Rapaport (1996) The Art of Molecular Dynamics Simulation.
ISBN 0-521-44561-2.
M. Griebel; S. Knapek; G. Zumbusch (2007). Numerical Simulation in Molecular Dynamics. Berlin, Heidelberg: Springer. ISBN 978-3-540-68094-9.
Frenkel, Daan; Smit, Berend (2002) [2001]. Understanding Molecular Simulation : from algorithms to applications. San Diego: Academic Press. ISBN 978-0-12-267351-1.
- J. M. Haile (2001) Molecular Dynamics Simulation: Elementary Methods.
ISBN 0-471-18439-X
- R. J. Sadus, Molecular Simulation of Fluids: Theory, Algorithms and Object-Orientation, 2002,
ISBN 0-444-51082-6
- Oren M. Becker, Alexander D. Mackerell, Jr., Benoît Roux, Masakatsu Watanabe (2001) Computational Biochemistry and Biophysics. Marcel Dekker.
ISBN 0-8247-0455-X. - Andrew Leach (2001) Molecular Modelling: Principles and Applications. (2nd Edition) Prentice Hall.
ISBN 978-0-582-38210-7.
Tamar Schlick (2002) Molecular Modeling and Simulation. Springer.
ISBN 0-387-95404-X.
William Graham Hoover (1991) Computational Statistical Mechanics, Elsevier,
ISBN 0-444-88192-1.- D. J. Evans and G. P. Morriss (2008) Statistical Mechanics of Nonequilibrium Liquids, Second Edition, Cambridge University Press,
ISBN 978-0-521-85791-8.
Bou-Rabee, Nawaf (2014). "Time Integrators for Molecular Dynamics". Entropy. 16 (1): 138–162. Bibcode:2013Entrp..16..138B. doi:10.3390/e16010138.
External links
| Wikimedia Commons has media related to Molecular dynamics. |
The GPUGRID.net Project (GPUGRID.net)
The Blue Gene Project (IBM)JawBreakers.org- Materials modelling and computer simulation codes
- A few tips on molecular dynamics
- Movie of MD simulation of water (Youtube)
- Live molecular dynamics simulation rendered at 1 frame per second


