Russulaceae
| Russulaceae | |
|---|---|
 | |
Russula emetica | |
Scientific classification | |
| Kingdom: | Fungi |
| Division: | Basidiomycota |
| Class: | Agaricomycetes |
| Order: | Russulales |
| Family: | Russulaceae Lotsy (1907)[1] |
Type genus | |
Russula Pers. (1796) | |
Genera | |
Boidinia | |
Synonyms[2][3] | |
| |
The Russulaceae are a diverse family of fungi in the order Russulales, with roughly 1,900 known species and a worldwide distribution. They comprise the brittlegills and the milk-caps, well-known mushroom-forming fungi that include some edible species. These gilled mushrooms are characterised by the brittle flesh of their fruitbodies.
In addition to these typical agaricoid forms, the family contains species with fruitbodies that are laterally stiped (pleurotoid), closed (secotioid or gasteroid), or crust-like (corticioid). Molecular phylogenetics has demonstrated close affinities between species with very different fruitbody types and has discovered new, distinct lineages.
An important group of root-symbiotic ectomycorrhizal fungi in forests and shrublands around the world includes Lactifluus, Multifurca, Russula, and Lactarius. The crust-forming genera Boidinia, Gloeopeniophorella, and Pseudoxenasma, all wood-decay fungi, have basal positions in the family.
Contents
1 Systematics and taxonomy
1.1 Placement of the family
1.2 Internal systematics
1.3 Species diversity
2 Description
2.1 Macroscopic characteristics
2.2 Microscopic characteristics
2.3 Genera distinction
3 Distribution
4 Ecology
4.1 Ectomycorrhizal symbiosis
4.2 Other types of mycorrhiza
4.3 Wood decay species
4.4 Hypogeous fruiting
4.5 Parasites
4.6 Threats and conservation
5 Edibility
6 Chemistry
7 References
8 External links
Systematics and taxonomy
The family Russulaceae was first validly named in 1907 by Dutch botanist Johannes Paulus Lotsy,[4] who included three genera: Russula, Lactarius, and Russulina (now considered a synonym of Russula). He emphasised features such as the granular flesh, thick gills, spiny spores, and milky hyphae and rounded cells (sphaerocytes).[1] A prior usage of "Russulariées" by French mycologist Ernst Roze in 1876[5] is not considered a valid publication, since the proper Latin termination for the family rank specified in article 18.4 of the nomenclature code was not used.[4][6]
Synonyms of Russulaceae include: Ernst Albert Gäumann's Lactariaceae (1926), Fernand Moreau's Asterosporaceae (1953),[2] and David Pegler and Thomas Young's Elasmomycetaceae (1979). The latter family was proposed to contain species with statismosporic (non-forcibly discharged) and symmetric spores, including the gasteroid genera Elasmomyces, Gymnomyces, Martellia, and Zelleromyces. Calonge and Martín reduced the Elasmomycetaceae to synonymy with the Russulaceae when molecular analysis confirmed the close genetic relationship between the gasteroid and agaricoid genera.[3]
Placement of the family
Historically, the gilled mushrooms of the family Russulaceae were classified with other gilled species in the order Agaricales,[7] but microscopical studies of spore and fruitbody flesh features raised the possibility that they were more closely related with certain "lower fungi" presenting nongilled, crust-like fruitbodies.[2][8][9] The use of molecular phylogenetics confirmed that these morphologically diverse fungi form a distinct lineage, first termed the "russuloid clade"[10][11] and today classified as order Russulales in the class Agaricomycetes.[12] The family's sister group within the order appears to be the crust-like Gloeocystidiellaceae.[13]
Internal systematics
.mw-parser-output table.clade{border-spacing:0;margin:0;font-size:100%;line-height:100%;border-collapse:separate;width:auto}.mw-parser-output table.clade table.clade{width:100%}.mw-parser-output table.clade td{border:0;padding:0;vertical-align:middle;text-align:center}.mw-parser-output table.clade td.clade-label{width:0.8em;border:0;padding:0 0.2em;vertical-align:bottom;text-align:center}.mw-parser-output table.clade td.clade-slabel{border:0;padding:0 0.2em;vertical-align:top;text-align:center}.mw-parser-output table.clade td.clade-bar{vertical-align:middle;text-align:left;padding:0 0.5em}.mw-parser-output table.clade td.clade-leaf{border:0;padding:0;text-align:left;vertical-align:middle}.mw-parser-output table.clade td.clade-leafR{border:0;padding:0;text-align:right}
| |||||||||||||||||||||||||||||||||
| Phylogeny of the Russulaceae; dashed lines indicate uncertain placement.[10][13][14][15] |
A 2008 molecular phylogenetic study clarified the relationships among the mushroom-forming species of the family.[15] The authors demonstrated the existence of four distinct lineages of gilled mushrooms, which led to the description of Multifurca as a new genus separated from Russula[15] and the segregation of Lactifluus from Lactarius.[16][17]
Genera with closed fruitbodies within the family are form taxa instead of natural groups: Arcangeliella, Gastrolactarius, and Zelleromyces are phylogenetically part of Lactarius, while Cystangium, Elasmomyces, Gymnomyces, Macowanites, and Martellia belong to Russula.[3][16] Nevertheless, some of these genus names are still in use, as many of the concerned species have not yet formally been synonymised with Lactarius or Russula.[18]
The crust-like genera Boidinia, Gloeopeniophorella, and Pseudoxenasma, formerly placed in the Corticiaceae or Gloeocystidiellaceae, are now classified in the Russulaceae and basal to the clade of mushroom-forming species described above.[13][18] Studies have so far failed to clearly circumscribe and place these genera within the family.[10][13][14]Boidinia in its current extent is polyphyletic, with some species not falling into the Russulaceae.[10]
Species diversity
Altogether, the Russulaceae comprise around 1,900 accepted species.[18]Russula is by far the largest genus with c. 1100 species, Lactarius has c. 550, Lactifluus c. 120, Boidinia 13, Multifurca 6, Gloeopeniophorella 6, and Pseudoxenasma 1 species.[18][19] Closed-fruitbody species not yet synonymised with Lactarius or Russula (see above) account for some 150 species.[18]
New species in the Russulaceae continue to be described from various regions, such as the US,[20] Guyana,[21] Brazil,[22] Patagonia,[23] Togo,[24] Sri Lanka,[25] or Thailand.[25] It has been estimated that the real number of Russula species in North America alone (currently around 400 described) might be as high as 2000.[26]Cryptic species may increase true diversity: some morphologically well-defined species, especially in Lactifluus, have been shown to actually encompass several phylogenetic species.[27][28][29]
Description
Macroscopic characteristics
@media all and (max-width:720px){.mw-parser-output .tmulti>.thumbinner{width:100%!important;max-width:none!important}.mw-parser-output .tmulti .tsingle{float:none!important;max-width:none!important;width:100%!important;text-align:center}}

Lactifluus vellereus (Russulaceae)

Suillus variegatus (Suillaceae)
Three major types of fruitbodies occur in the Russulaceae: agaricoid and pleurotoid forms with a cap, gills, and a stipe; forms with closed (gasteroid) or partially closed (secotioid) fruitbodies, and corticioid, crust-like forms.
The agaricoid species in Lactarius, Lactifluus, Multifurca, and Russula are readily distinguished from other gilled mushrooms by the consistency of their flesh, which is granular, brittle and breaks easily, somewhat like a piece of chalk.[1][30] Russulaceae never have a volva,[30] but a partial veil can be found in some tropical species.[31][32] Gills are adnate to decurrent, and the colour of the spore print ranges from white to ochre or orange[15][33] (with the brown-spored Lactarius chromospermus as an exception[34]).
Caps can be dull to very colourful, the latter especially in Russula;[33] their size ranges from 17 mm diameter or less in Russula campinensis[35] to 30 cm (12 in) in Lactifluus vellereus.[33] Concentrically ringed (zonate) caps occur in all Multifurca[15] and several Lactarius species.[33] Laterally stiped (pleurotoid) fruitbodies exist in some, mainly tropical Lactifluus and Russula species.[35][36][37][38] Taste is a distinguishing characteristic in many species, from mild to very acrid.[33] A conspicuous feature of the "milk-caps" in Lactarius, Lactifluus, and Multifurca furcata is the latex or "milk" their fruitbodies exude when bruised.[15][30]
The secotioid and gasteroid species in Lactarius and Russula are derived from agaricoid forms.[15][39] Secotioid species still have a stipe but the cap does not open fully, while in gasteroid species, fruitbodies are completely closed and the stipe is reduced; in both cases, the spore-bearing structure is made up of convoluted gills that are more or less crowded and anastomosed.[39] These closed-fruitbody species represent a continuum of secotioid to gasteroid, above-ground to below-ground fruitbodies, with spores forcibly discharged or not.[25][39][40] Secotioid or gasteroid Lactarius exude latex just like their agaricoid relatives.[3][25][40]
The corticioid species of Boidinia, Gloeopeniophorella, and Pseudoxenasma develop crust-like fruitbodies with a smooth, porous, or flaky surface and grow on tree logs or dead branches.[10][41][42][43]
|
| *These species belong phylogenetically to Lactarius. |





Microscopic characteristics
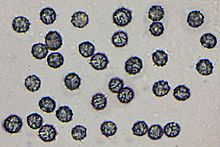
Spores of Lactarius rubidus, showing ornamentation with a blue, amyloid stain reaction that is typical for Russulaceae
All Russulaceae, including the corticioid species, are characterised by spherical to elliptic basidiospores with a faint to very distinct (e.g. warty, spiny, or crested) ornamentation that stains bluish-black with Melzer's reagent (an amyloid stain reaction).[10][30]Basidia (spore-bearing cells) are usually club-shaped and four-spored.[44] Russulaceae species do not have clamp connections.[45]
Characteristic cells with an oily content (gloeocystidia) are found in the hymenium. In Russulaceae, these show a positive colour reaction when treated with sulfoaldehydes (sulfovanillin is mostly used).[10][30] They are also present in the hyphal sheath of ectomycorrhizal roots colonised by Russulaceae.[14]
The feature responsible for the brittle fruitbody structure in the mushroom-forming species are globular cells, called sphaerocytes or sphaerocysts, that compose the flesh (trama) alongside the usual hyphae.[30] Sometimes, these cells are clustered, and the position and arrangement of these clusters differs among genera.[30]
Another particular trama cell type are lactiferous hyphae (also lactifers). These are hyphae carrying the "milk" or "latex" exuded by the milk-caps; they react positively with sulfoaldehydes, form an abundantly branched system in the trama and end as pseudocystidia in the hymenium.[30] In general, only Lactarius, Lactifluus and Multifurca furcata possess lactifers.[15] In Russula, similar hyphae can sometimes be observed in the trama, but these are not as abundantly branched as real lactifers and do not extend into the hymenium as pseudocystidia.[30] This traditional distinction line between the "milk-caps" and Russula is however less evident in some tropical species presenting intermediate states.[36]
Genera distinction


Some characteristics of the mushroom-forming genera (marked with * below) can be less obvious or absent in tropical species.[15][36] Distinguishing between Lactarius and Lactifluus based on morphology alone is quite difficult, as clear synapomorphies for both genera have yet to be identified.[17] Most field guides treat the two genera together, often because Lactifluus is not yet recognised as a separate genus.[33][46]
Boidinia: corticioid; loose texture; surface smooth, with pores, or flaky; spores spherical with spiny to warty ornamentation.[10][41] Note that the genus is polyphyletic and needs to be redefined.[10]
Gloeopeniophorella: corticioid; surface almost smooth; hyphae without clamp connections; thick-walled cystidia (metuloids) and gloeocystidia present; spores with wrinkled (rugose) ornamentation.[43]
Lactarius: agaricoid or gasteroid; exuding latex*; caps sometimes zonate, viscose or glutinate, but never annulate; rarely thick-walled cells in cuticles of the cap (pileipellis) and the stipe (stipitipellis) and sphaerocytes in the gills.[15][17]
Lactifluus: agaricoid or pleurotoid; exuding latex*; caps never zonate, viscose or glutinate, but sometimes annulate; thick-walled cells in cap and stipe cuticles; often sphaerocytes in the gill trama.[15][17]
Multifurca: agaricoid; caps zonate (also visible in cut through trama); gills regularly forked; only M. furcata exuding latex; spore print orange; spores very small; microscopical trama and hymenium features very variable.[15]
Pseudoxenasma: corticioid; wax-like texture; gloeocystidia with spherical apical appendices; basidia developing laterally on hyphae (pleurobasidia); spores broadly ellipsoid to roughly spherical, with warty ornamentation.[42]
Russula: agaricoid, gasteroid or pleurotoid; never exuding latex; caps often brightly coloured with stipe and gills much paler; caps not zonate*; spore print white, cream, ochre, or orange; no true lactiferous hyphae*; sphaerocytes abundant in gill, cap, and stipe trama.[15][30]
Distribution
The Russulaceae as a whole have a worldwide distribution, but patterns differ among genera. Russula is the most widespread, found in North,[47][48] Central[49][50] and South America,[32][51] Europe,[33] temperate[52][53] and tropical Asia,[54][55] Africa,[56] and Australasia.[36][57][58] It is the only Russulaceae genus that occurs in the Nothofagus zone of temperate South America.[59]
Lactarius is mainly known from the north temperate zone, but some species also occur in tropical Asia and Africa.[17]Lactifluus has a more tropical distribution than Lactarius, with most species known from tropical Africa, Asia, South America, and Australasia, but some also occurring in the north temperate zone.[17]Multifurca is the rarest among the four mushroom genera, known only from some punctual records in North and Central America, Asia, and Australasia.[15][19]
Species of Lactarius, Lactifluus, and Russula have repeatedly been introduced with trees outside their native range: An overview article lists introductions in Chile, Argentina, Uruguay, Brazil, the USA, Great Britain, the Faroe Islands, South Africa, China, Thailand, and New Zealand.[60]
Among the corticioid genera, Pseudoxenasma is only known from Europe.[61] In contrast, Boidinia species have been found in Europe,[62] Taiwan,[63] and Japan,[41] and Gloeopeniophorella species in North America,[64] South America,[43][65] Europe,[66] West Africa,[65] Taiwan,[63] Australia,[43] and New Zealand.[43]
Ecology
Ectomycorrhizal symbiosis


Miombo woodland, Malawi
The genera Lactarius, Lactifluus, Multifurca and Russula form a mutualistic ectomycorrhizal root symbiosis with trees and shrubs, exchanging mineral nutrients for photosynthetic sugar. They are one of several fungal lineages that have evolved such a lifestyle and are sometimes referred to as the "/russula-lactarius" clade in the scientific literature.[67] Worldwide, they are one of the most frequently encountered lineages on ectomycorrhizal roots.[68] While some tropical species were initially believed to be parasitic, the observation that species fruiting on tree trunks do form ectomycorrhiza in tropical Guyana supports the view of an exclusively symbiotic lineage.[35]
Associations are known with several plant families. In the Northern Hemisphere, these are essentially the well-known ectomycorrhizal trees and shrubs in the Betulaceae, Fagaceae, Pinaceae and Salicaceae,[33][46] but in arctic and alpine habitats, Russulaceae also associate with Bistorta vivipara (Polygonaceae),[69]Kobresia (Cyperaceae),[70] and Dryas octopetala (Rosaceae),[71] ectomycorrhizal plants untypic in their respective families. In the tropics, known plant partners include Dipterocarpaceae,[55][72]Fabaceae,[35][72]Nyctaginaceae,[73][74]Phyllanthaceae,[72] Polygonaceae (Coccoloba),[74]Sarcolaenaceae,[75] and the gymnosperm Gnetum gnemon,[76] and in the Southern Hemisphere, Nothofagaceae,[58][59][77] Myrtaceae (Eucalyptus[57] and Leptospermum[58][77]), and Rhamnaceae (Pomaderris).[78] Some Russulaceae are quite specialised in their ectomycorrhizal symbiosis, such as Lactarius and Russula species that only grow with Cistus shrubs in the Mediterranean basin.[79]
The different plant partners are reflected in the wide variety of habitats worldwide.[80] Ectomycorrhizal Russulaceae have been observed in arctic and alpine tundra,[69]boreal and alpine forest,[52][81]north temperate forest,[33][46]mires,[82] mediterranean forests and scrub (maquis),[79][83]miombo woodland,[56]tropical lowland rainforest,[55][74]tropical cloud forest,[84]tropical dry forest,[85] Australian eucalypt woodlands,[86] and south temperate forests.[59][77][78] Where they are introduced, they typically grow in plantations of their native host species, e.g. with pine in South Africa,[87]Eucalyptus in Thailand,[88] or birch in New Zealand.[77]
Other types of mycorrhiza

Root tips of the myco-heterotrophic plant Monotropa uniflora, with a mycorrhizal sheath formed by Russula brevipes
Some of the ectomycorrhizal Russulaceae are also involved in other types of root symbioses with plants.
A mutualistic association similar to ectomycorrhiza but with some hyphae penetrating into the plant root cells, termed arbutoid mycorrhiza,[89] is formed by Russulaceae with shrubs of the genera Arbutus[83] and Arctostaphylos,[90] both in subfamily Arbutoideae of the Ericaceae.[83][90]
Some Russulaceae are associated with myco-heterotrophic plants of the Ericaceae subfamily Monotropoideae, forming monotropoid mycorrhiza.[91][92] This is an epiparasitic relationship, where the heterotrophic plant ultimately derives its carbon from the primary, ectomycorrhizal plant partner of the fungus.[92] The association is often very specific, with the heterotrophic plants only associating with selected fungus partners, including Russulaceae.[91][93]
Russulaceae are also an important group of orchid mycorrhizal fungi.[94] This symbiosis is mutualistic in the case of green orchids,[89] but a partly or fully epiparasitic relationship in the case of myco-heterotrophic[95][96] and mixotrophic[97] orchids, respectively. In some cases, the association with Russulaceae is, as in monotropoid mycorrhiza, very specific: the Mediterranean orchid Limodorum abortivum predominantly associates with Russula delica and closely related species;[97] in Corallorhiza maculata, different genotypes of the same species have distinct Russula partners.[95]
Wood decay species
The corticioid species in Boidinia, Gloeopeniophorella, and Pseudoxenasma are saprotrophic, wood-degrading fungi that develop on dead wood.[10] Their early-branching positions in the phylogeny suggests this has been the ancestral trophic mode of the Russulaceae, and that the mycorrhizal lifestyle (see above) evolved later.[10] The saprotrophic nature of these species has been questioned, based on the observation that other inconspicuous, crust-forming fungi are ectomycorrhizal;[14] a subsequent author reaffirms nevertheless that "[n]one of the corticioid species in the family shows any sign of mycorrhizal activity."[13]
Hypogeous fruiting
Hypogeous fruitbodies, or fruitbodies developing below ground, occur in Lactarius and Russula and have previously been considered as distinct genera (see Systematics and taxonomy: Internal systematics). As such species are especially diverse in some warm and dry regions, e.g. in Spain,[3] California,[98] or Australia,[57] below-ground fruiting has been interpreted as an adaptation to drought.[39] However, hypogeous Russulaceae are also known from cold temperate regions[23][99] and tropical rainforest.[25] The fact that hypogeous species in the Russulaceae do not form their own lineages but are scattered in Russula or Lactarius shows that this type of fruiting evolved several times.[25] It is believed that these changes are evolutionarily quite recent.[25]
Parasites

Asterophora parasitica grows on rotting Russulaceae fruitbodies.
Russulaceae fruitbodies are subject to parasitisation by other fungi. The genus Asterophora develops on old fruitbodies of the mushroom species in the family,[46] as does Dendrocollybia racemosa on at least Russula crassotunicata.[100] Fruitbodies of Lactifluus or Russula species otherwise hot-tasting and unpalatable are regarded as choice edibles in North America when infected by the "lobster mushroom" Hypomyces lactifluorum.[46] Heterotrophic plants, including orchids or monotropoids, also parasitise ectomycorrhizal Russulaceae and their plant partners – see above, Other types of mycorrhiza.
Threats and conservation
As with most fungi,[101] little information is available on the threat of extinction for Russulaceae species, and they have not been assessed in the International Union for the Conservation of Nature's Red List.[102] However, national lists contain some species of Lactarius, Lactifluus and Russula, indicating that they have small populations and are endangered, e.g. in Great Britain,[103] Switzerland,[104] the Czech Republic,[105] and New Zealand.[106]
Although data on Russulaceae themselves are scarce, more is known about the habitats they occur in, especially for the ectomycorrhizal species which depend on their host plants: Several of these habitats are affected by loss or degradation, such as peatlands,[107] Mediterranean forests and scrub[108] or tropical African dry woodland.[109] Similarly, dead wood, the habitat of the corticioid Russulaceae, is rare in many exploited forests and needs special management.[110]
Recent studies have found some traditional Russulaceae species to comprise several cryptic species (see Systematics and taxonomy: Species diversity). This may imply that distribution range and population size for each of such distinct species are smaller than previously thought.[111]
Edibility

Lactarius indigo and another Lactarius species for sale on a market in Guatemala
Several species of Lactarius, Lactifluus and Russula are valued as excellent edible mushrooms. This is the case for example for the north temperate species Lactarius deliciosus, Lactifluus volemus, or Russula vesca, and other species are popular in other parts of the world, e.g. Lactarius indigo in Mexico, or Lactifluus edulis in tropical Africa.[112] Some species, like Russula vesca, can even be eaten raw.[113] The brittle texture of Russula fruitbodies makes them different from other mushrooms and is not appreciated by some.[114]
Several species have a hot to very acrid taste and can cause gastrointestinal symptoms.[115] Despite this, such species are eaten in some regions, e.g. Lactarius torminosus in Finland[116] or Russia.[117] Often, they are parboiled or pickled to make them palatable,[118] and sometimes, they are used as spice, for example Russula emetica in Eastern Europe.[119] Some species are however truly poisonous: the East Asian and North American Russula subnigricans causes rhabdomyolysis and is potentially lethal,[120] and Lactarius turpis from Eurasia contains a mutagenic substance.[121]
Cultivation of edible Russulaceae, as in other ectomycorrhizal fungi, is challenging, since the presence of host trees is required. In spite of this difficulty, the European Lactarius deliciosus has been successfully grown in "mushroom orchards" in New Zealand.[122]
Chemistry

Lactarius indigo contains a striking blue pigment.
Fruitbodies of Russulaceae have been the subject of natural product research, and different classes of organic compounds have been isolated from them.
Aroma compounds are responsible for the particular odour or taste in some species, e.g. sotolon in the fenugreek-smelling Lactarius helvus,[123] or the similar quabalactone III in Lactarius rubidus which causes a maple syrup-like odour in dried specimens.[124]Pigments have been isolated from brightly coloured species, e.g. (7-isopropenyl-4-methylazulen-1-yl)methyl stearate from the blue Lactarius indigo[125] or russulaflavidin and a derivative from the yellow Russula flavida.[126] Some Russula species contain pigmented pteridine derivatives called russupteridines that are not found in the milk-caps.[127]Sesquiterpenes are characteristic secondary metabolites of many Russulaceae, especially milk-caps which have been quite intensively studied.[128][129] They are thought to be responsible for the hot taste in many species and may have deterrent, antifeeding functions in nature.[128]
Other metabolites isolated from different species include dibenzonaphtyridinone alkaloids,[128]prenylated phenols,[128]benzofurans,[128]chromenes,[128] natural rubber (polyisoprene),[130]sterols,[131] and the sugar alcohol volemitol.[132] Among toxic substances, Lactarius turpis contains the mutagenic alkaloid necatorin,[121] and the small compound cycloprop-2-ene carboxylic acid has been identified as the toxic agent in Russula subnigricans.[133] Some secondary metabolites showed antibiotic properties in laboratory tests.[128] An ethanolic extract of Russula delica was antibacterial,[134] and a lectin from Russula rosea showed antitumor activity.[135]
References
^ abc Lotsy JP (1907). Vorträge über botanische Stammesgeschichte [Lectures on botanical phylogeny, held at the Imperial University of Leiden] (in German). Jena, Germany: Gustav Fischer. p. 708..mw-parser-output cite.citation{font-style:inherit}.mw-parser-output q{quotes:"""""""'""'"}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}

^ abc Pegler DN, Young TW (1979). "The gasteroid Russulales". Transactions of the British Mycological Society. 72 (3): 353–388. doi:10.1016/S0007-1536(79)80143-6.
^ abcde Calonge FD, Martín MP (2000). "Morphological and molecular data on the taxonomy of Gymnomyces, Martellia and Zelleromyces (Elasmomycetaceae, Russulales)" (PDF). Mycotaxon. 76: 9–15.
^ ab "Russulaceae Lotsy, Truffe, ed. 2: 708 (1907)". MycoBank. International Mycological Association. Retrieved 2014-12-28.
^ Roze ME (1876). "Catalogue des agaricinées observées aux environs de Paris" [Catalogue of agarics observed around Paris]. Bulletin de la Société botanique de France (in French). 23 (3): 108–115 (see p. , 110). doi:10.1080/00378941.1876.10825634.
^ McNeill J, Barrie FR, Buck WR, Demoulin V, eds. (2012). "Article 18.4". International Code of Nomenclature for algae, fungi, and plants (Melbourne Code), adopted by the Eighteenth International Botanical Congress Melbourne, Australia, July 2011 (electronic ed.). Bratislava: International Association for Plant Taxonomy.
^ Singer R (1986). The Agaricales in Modern Taxonomy (4th ed.). Königstein im Taunus, Germany: Koeltz Scientific Books. ISBN 978-3-87429-254-2.
^ Donk MA (1971). "Progress in the study of the classification of the higher Basidiomycetes". In Petersen RH. Evolution in the Higher Basidiomycetes. Knoxville. pp. 3–25. OCLC 139485.
^ Oberwinkler F (1977). "Das neue System der Basidiomyceten" [The new basidiomycete system]. In Frey W, Hurka H, Oberwinkler F. Beiträge zur Biologie der niederen Pflanzen (in German). Stuttgart; New York: Gustav Fischer Verlag. pp. 59–104. ISBN 978-3-437-30262-6.
^ abcdefghijk Larsson E, Larsson KH (2003). "Phylogenetic relationships of russuloid basidiomycetes with emphasis on aphyllophoralean taxa". Mycologia. 95 (6): 1037–65. doi:10.2307/3761912. JSTOR 3761912. PMID 21149013.
^ Hibbett DH, Thorn RG (2001). "Basidiomycota: Homobasidiomycetes". In McLaughlin DJ, McLaughlin EG, Lemke PA. The Mycota. VIIB. Systematics and Evolution. Berlin: Springer-Verlag. pp. 121–168. ISBN 978-3-540-58008-9.
^ Hibbett DS, Binder M, Bischoff JF, Blackwell M, Cannon PF, Eriksson OE, et al. (May 2007). "A higher-level phylogenetic classification of the Fungi" (PDF). Mycological Research. 111 (Pt 5): 509–47. doi:10.1016/j.mycres.2007.03.004. PMID 17572334. Archived from the original (PDF) on 2009-03-26.
^ abcde Larsson KH (September 2007). "Re-thinking the classification of corticioid fungi". Mycological Research. 111 (Pt 9): 1040–63. doi:10.1016/j.mycres.2007.08.001. PMID 17981020.
^ abcd Miller SL, Larsson E, Larsson KH, Verbeken A, Nuytinck J (2006). "Perspectives in the new Russulales" (PDF). Mycologia. 98 (6): 960–70. doi:10.3852/mycologia.98.6.960. PMID 17486972.
^ abcdefghijklmn Buyck B, Hofstetter V, Eberhardt U, Verbeken A, Kauff F (2008). "Walking the thin line between Russula and Lactarius: The dilemma of Russula sect. Ochricompactae" (PDF). Fungal Diversity. 28: 15–40.
^ ab Buyck B, Hofstetter V, Verbeken A, Walleyn R (2010). "Proposal to conserve Lactarius nom. cons. (Basidiomycota) with conserved type". Taxon. 59: 447–453.
^ abcdef Verbeken A, Nuytinck J (2013). "Not every milkcap is a Lactarius" (PDF). Scripta Botanica Belgica. 51: 162–168.
^ abcde Kirk PM. "Species Fungorum (version October 2014). In: Species 2000 & ITIS Catalogue of Life". Species 2000 & ITIS. Retrieved 2014-10-30.
^ ab Lebel T, Dunk CW, May TW (2013). "Rediscovery of Multifurca stenophylla (Berk.) T.Lebel, C.W.Dunk & T.W.May comb. nov. (Russulaceae) from Australia". Mycological Progress. 12 (3): 497–504. doi:10.1007/s11557-012-0856-4.
^ Arora D, Nguyen N (2014). "A new species of Russula, subgenus Compactae from California" (PDF). North American Fungi. 8 (8): 1–7. ISSN 1937-786X.
^ Miller S, Aime MC, Henkel TW (2013). "Russulaceae of the Pakaraima Mountains of Guyana 2. New species of Russula and Lactifluus" (PDF). Mycotaxon. 121 (1): 233–253. doi:10.5248/121.233.
^ Sá MC, Wartchow F (2013). "Lactifluus aurantiorugosus (Russulaceae), a new species from Southern Brazil" (PDF). Darwiniana, nueva serie. 1: 54–60.
^ ab Trierveiler-Pereira L, Smith ME, Trappe JM, Nouhra ER (2014). "Sequestrate fungi from Patagonian Nothofagus forests: Cystangium (Russulaceae, Basidiomycota)". Mycologia. 107 (1): 90–103. doi:10.3852/13-302. PMID 25232070.
^ Maba DL, Guelly AK, Yorou NS, De Kesel A, Verbeken A, Agerer R (June 2014). "The genus Lactarius s. str. (Basidiomycota, Russulales) in Togo (West Africa): phylogeny and a new species described". IMA Fungus. 5 (1): 39–49. doi:10.5598/imafungus.2014.05.01.05. PMC 4107895. PMID 25083405.
^ abcdefg Verbeken A, Stubbe D, van de Putte K, Eberhardt U, Nuytinck J (June 2014). "Tales of the unexpected: angiocarpous representatives of the Russulaceae in tropical South East Asia". Persoonia. 32 (1): 13–24. doi:10.3767/003158514X679119. PMC 4150074. PMID 25264381.
^ Vellinga EC (2013). "The North American Mycoflora Project: Without a sequenced specimen it is a rumor". McIlvainea (22): V22/north_american_mycoflora.html.
^ Van de Putte K, Nuytinck J, Stubbe D, Le HT, Verbeken A (2010). "Lactarius volemus sensu lato (Russulales) from northern Thailand: Morphological and phylogenetic species concepts explored". Fungal Diversity. 45 (1): 99–130. doi:10.1007/s13225-010-0070-0.
^ De Crop E, Nuytinck J, Van de Putte K, Lecomte M, Eberhardt U, Verbeken A (2014). "Lactifluus piperatus (Russulales, Basidiomycota) and allied species in Western Europe and a preliminary overview of the group worldwide". Mycological Progress. 13 (3): 493–511. doi:10.1007/s11557-013-0931-5.
^ "Contrasting evolutionary patterns in two sister genera of macrofungi: Lactarius and Lactifluus". Ghent University. Archived from the original on 2014-10-06. Retrieved 2014-10-10.
^ abcdefghij "Russulales News / Characteristics of the russuloid fungi". Museo delle scienze. Retrieved 2014-11-01.
^ Heim R (1938). "Diagnoses latines d'espèces et variétés nouvelles de Lactario-russulés du domaine oriental de Madagascar [Latin diagnoses for new species and varieties of Lactario-Russulas from the Eastern domain of Madagascar]". Candollea (in French and Latin). 7: 374–393.
^ ab Singer R, Aurajuo I, Ivory MH (1983). "The ectotrophically mycorrhizal fungi of the neotropical lowlands, especially central Amazonia". Beihefte zur Nova Hedwigia: 1–352.
^ abcdefghi Courtecuisse R, Duhem B (2013). Champignons de France et d'Europe [Mushrooms of France and Europe]. Guide Delachaux (in French). Paris: Delachaux & Niestlé. ISBN 978-2-603-02038-8.
^ Buyck B, Verbeken A (1995). "Studies in tropical African Lactarius species, 2: Lactarius chromospermus Pegler". Mycotaxon. 56: 427–442.
^ abcd Henkel T, Aime MC, Miller SL (2000). "Systematics of pleurotoid Russulaceae from Guyana and Japan, with notes on their ectomycorrhizal status". Mycologia. 92 (6): 1119–1132. doi:10.2307/3761479. JSTOR 3761479.
^ abcd Buyck B, Horak E (1999). "New taxa of pleurotoid Russulaceae". Mycologia. 91 (3): 532–537. doi:10.2307/3761355. JSTOR 3761355.
^ Wang XH, Stubbe D, Verbeken A (2012). "Lactifluus parvigerardii sp. nov., a new link towards the pleurotoid habit in Lactifluus subgen. Gerardii (Russulaceae, Russulales)". Cryptogamie, Mycologie. 332 (2): 181–190. doi:10.7872/crym.v33.iss2.2012.181.
^ Morozova OV, Popov ES, Kovalenko AE (2013). "Studies on mycobiota of Vietnam. II. Two new species of Lactifluus (Russulaceae) with pleurotoid basidiomata" (PDF). Mikologia i Fitopatologia. 47 (2): 92–102.
^ abcd Thiers HD (1984). "The secotioid syndrome" (PDF). Mycologia. 76 (1): 1–8. doi:10.2307/3792830. JSTOR 3792830.
^ ab Desjardin DE (2003). "A unique ballistosporic hypogeous sequestrate Lactarius from California". Mycologia. 95 (1): 148–155. doi:10.2307/3761974. JSTOR 3761974.
^ abc Maekawa N (1994). "Taxonomic study of Japanese Corticiaceae (Aphyllophoraceae) II". Report of the Tottori Mycological Institute. 32: 1–123.
^ ab Hjortstam K, Larsson KH (1976). "Pseudoxenasma, a new genus of Corticiaceae (Basidiomycetes)". Mycotaxon. 4 (1): 307–311.
^ abcde Hjortstam K, Ryvarden L (2007). "Studies in corticioid fungi from Venezuela III (Basidiomycotina, Aphyllophorales)". Synopsis Fungorum. 23: 56–107.
^ Cannon PF, Kirk PM (2007). Fungal Families of the World. Wallingford, UK: CAB International. pp. 317–318. ISBN 978-0-85199-827-5.
^ Ammirati JF, Traquair JA, Horgen PA (1985). Poisonous Mushrooms of Canada: Including other Inedible Fungi. Markham, Ontario: Fitzhenry & Whiteside in cooperation with Agriculture Canada and the Canadian Government Publishing Centre, Supply and Services Canada. p. 57. ISBN 978-0-88902-977-4.
^ abcde Bessette AE, Bessette AR, Fischer D (1996). Mushrooms of Northeastern North America. Syracuse, New York: Syracuse University Press. ISBN 978-0-8156-0388-7.
^ Earle FS (1902). "A key to the North American species of Russula – I". Torreya. 2 (7): 101–103. JSTOR 40594086.
^ Earle FS (1902). "A key to the North American species of Russula – II". Torreya. 2 (8): 117–119. JSTOR 40594096.
^ Gómez-Pignataro LD, Alfaro RM (1996). "Basidiomicetes de Costa Rica. De Russulae novae". Revista de Biología Tropical (in Spanish and English). 44 (Suppl. 4): 25–37.
^ Buyck B, Ovrebo CL (2002). "New and interesting Russula species from Panama". Mycologia. 94 (5): 888–901. doi:10.2307/3761704. JSTOR 3761704. PMID 21156563.
^ Sá MC, Baseia IG, Wartchow F (2013). "Checklist of Russulaceae from Brazil" (PDF). Mycotaxon. online 125: 303.
^ ab Guo J, Karunarathna SC, Mortimer PE, Xu J, Hyde KD (2014). "Phylogenetic diversity of Russula from Xiaozhongdian, Yunnan, China, inferred from Internal Transcribed Spacer sequence data" (PDF). Chiang Mai Journal of Science. 41 (4): 811–821.
[permanent dead link]
^ Gorbunova IA (2014). "Biota of agaricoid and gasteriod basidiomycetes of dryad tundras of the Altai-Sayan mountain area (Southern Siberia)". Contemporary problems of Ecology. 7 (1): 39–44. doi:10.1134/S1995425514010065.
^ Lee LS, Alexander IJ, Watling R (1997). "Ectomycorrhizas and putative ectomycorrhizal fungi of Shorea leprosula Miq. (Dipterocarpaceae)". Mycorrhiza. 7 (2): 63–81. doi:10.1007/s005720050165.
^ abc Natarajan KN, Senthilarasu G, Kumaresan V, Rivière T (2005). "Diversity in ectomycorrhizal fungi of a dipterocarp forest in Western Ghats" (PDF). Current Science. 88 (12): 1893–1895.
^ ab Verbeken A, Buyck B (2002). "Diversity and ecology of tropical ectomycorrhizal fungi in Africa". In Watling R, Frankland JC, Ainsworth AM, Isaac S, Robinson CH. Tropical Mycology: Macromycetes (PDF). Wallingford, UK: CAB International. pp. 11–21. Archived from the original (PDF) on 2014-10-06.
^ abc Bougher NL (1996). "Diversity of ectomycorrhizal fungi associated with eucalypts in Australia". In Brundett M, Dell B, Malajczuk N, Mingqin G. Mycorrhizas for plantation forestry in Asia (PDF). ACIAR Proceedings. pp. 8–15. ISBN 978-1-86320-167-4.
^ abc McNabb RF (1973). "Russulaceae of New Zealand 2. Russula Pers. ex S. F. Gray". New Zealand Journal of Botany. 11 (4): 673–730. doi:10.1080/0028825X.1973.10430308.
^ abc Singer R (1953). "Four years of mycological work in southern South America". Mycologia. 45 (6): 865–891. JSTOR 4547770.
^ Vellinga EC, Wolfe BE, Pringle A (March 2009). "Global patterns of ectomycorrhizal introductions". The New Phytologist. 181 (4): 960–73. doi:10.1111/j.1469-8137.2008.02728.x. PMID 19170899.
^ "Pseudoxenasma verrucisporum". Global Biodiversity Information Facility (GBIF). Retrieved 2014-11-20.
^ Bernicchia A, Gorjón SP (2010). Corticiaceae s.l. Fungi Europaei. 12. Alassio, Italy: Edizioni Candusso. ISBN 978-88-901057-9-1.
^ ab Wu SH (1996). "Studies on Gloeocystidiellum sensu lato (Basidiomycotina) in Taiwan". Mycotaxon. 58: 1–68.
^ Ginns J, Freeman GW (1994). "The Gloeocystidiellaceae (Basidiomycota, Hericiales) of North America". Bibliotheca Mycologica. 157: 1–118.
^ ab Boidin J, Lanquetin P, Gilles G (1997). "Le genre Gloeocystidiellum sensu lato (Basidiomycotina)". Bulletin de la Société Mycologique de France (in French). 113 (1): 1–80.
^ Eriksson J, Ryvarden L (1975). The Corticiaceae of North Europe. 3. Oslo: Fungiflora.
^ "UNITE database: EcM genera and lineages". University of Tartu. Archived from the original on 2014-10-25. Retrieved 2014-10-25.
^ Tedersoo L, Nara K (January 2010). "General latitudinal gradient of biodiversity is reversed in ectomycorrhizal fungi". The New Phytologist. 185 (2): 351–4. doi:10.1111/j.1469-8137.2009.03134.x. PMID 20088976.
^ ab Gardes M, Dahlberg A (1996). "Mycorrhizal diversity in arctic and alpine tundra: An open question". New Phytologist. 133 (1): 147–157. doi:10.1111/j.1469-8137.1996.tb04350.x.
^ Gao Q, Yang ZL (April 2010). "Ectomycorrhizal fungi associated with two species of Kobresia in an alpine meadow in the eastern Himalaya". Mycorrhiza. 20 (4): 281–7. doi:10.1007/s00572-009-0287-5. PMC 2845889. PMID 20012655.
^ Harrington TJ, Mitchell DT (2002). "Characterization of Dryas octopetala ectomycorrhizas from limestone karst vegetation, western Ireland". Canadian Journal of Botany. 80 (9): 970–982. doi:10.1139/b02-082.
^ abc Bâ A, Duponnois R, Diabaté, Dreyfus B (2011). Les champignons ectomycorhiziens des arbres forestiers en Afrique de l’Ouest [The ectomycorrhizal fungi of planted trees in West Africa] (in French). Marseille: Institut de recherche pour le développement (IRD). pp. 88–140. ISBN 978-2-7099-1684-4.
^ Haug I, Weiss M, Homeier J, Oberwinkler F, Kottke I (March 2005). "Russulaceae and Thelephoraceae form ectomycorrhizas with members of the Nyctaginaceae (Caryophyllales) in the tropical mountain rain forest of southern Ecuador". The New Phytologist. 165 (3): 923–36. doi:10.1111/j.1469-8137.2004.01284.x. PMID 15720703.
^ abc Tedersoo L, Sadam A, Zambrano M, Valencia R, Bahram M (April 2010). "Low diversity and high host preference of ectomycorrhizal fungi in western Amazonia, a neotropical biodiversity hotspot". The ISME Journal. 4 (4): 465–71. doi:10.1038/ismej.2009.131. PMID 19956273.
^ Ducousso M, Ramanankierana H, Duponnois R, Rabévohitra R, Randrihasipara L, Vincelette M, Dreyfus B, Prin Y (2008). "Mycorrhizal status of native trees and shrubs from eastern Madagascar littoral forests with special emphasis on one new ectomycorrhizal endemic family, the Asteropeiaceae". The New Phytologist. 178 (2): 233–8. doi:10.1111/j.1469-8137.2008.02389.x. PMID 18371004.
^ Tedersoo L, Põlme S (November 2012). "Infrageneric variation in partner specificity: multiple ectomycorrhizal symbionts associate with Gnetum gnemon (Gnetophyta) in Papua New Guinea". Mycorrhiza. 22 (8): 663–8. doi:10.1007/s00572-012-0458-7. PMID 22892664.
^ abcd McNabb RF (1971). "The Russulaceae of New Zealand 1. Lactarius DC ex S. F. Gray". New Zealand Journal of Botany. 9 (1): 46–66. doi:10.1080/0028825X.1971.10430170.
^ ab Tedersoo L, Jairus T, Horton BM, Abarenkov K, Suvi T, Saar I, Kõljalg U (2008). "Strong host preference of ectomycorrhizal fungi in a Tasmanian wet sclerophyll forest as revealed by DNA barcoding and taxon-specific primers". The New Phytologist. 180 (2): 479–90. doi:10.1111/j.1469-8137.2008.02561.x. PMID 18631297.
^ ab Comandini O, Contu M, Rinaldi AC (September 2006). "An overview of Cistus ectomycorrhizal fungi" (PDF). Mycorrhiza. 16 (6): 381–395. doi:10.1007/s00572-006-0047-8. PMID 16896800. Archived from the original (PDF) on 2016-03-04.
^ "Russulales News / Where do we find Russulales?". Museo delle scienze. Retrieved 2014-11-01.
^ Toljander JF, Eberhardt U, Toljander YK, Paul LR, Taylor AF (2006). "Species composition of an ectomycorrhizal fungal community along a local nutrient gradient in a boreal forest". The New Phytologist. 170 (4): 873–83. doi:10.1111/j.1469-8137.2006.01718.x. PMID 16684245.
^ Thormann MN, Rice AV (2007). "Fungi from peatlands" (PDF). Fungal Diversity. 24: 241–299.
^ abc Richard F, Millot S, Gardes M, Selosse MA (June 2005). "Diversity and specificity of ectomycorrhizal fungi retrieved from an old-growth Mediterranean forest dominated by Quercus ilex". The New Phytologist. 166 (3): 1011–23. doi:10.1111/j.1469-8137.2005.01382.x. PMID 15869659.
^ Halling RE, Mueller GM. "Agarics and boletes of neotropical oakwoods". In Watling R, Frankland JC, Ainsworth AM, Isaac S, Robinson CH. Tropical Mycology: Macromycetes (PDF). Wallingford, UK: CAB International. pp. 1–10. Archived from the original (PDF) on 2014-10-07.
^ Phosri C, Põlme S, Taylor AF, Kõljalg U, Suwannasai N, Tedersoo L (2012). "Diversity and community composition of ectomycorrhizal fungi in a dry deciduous dipterocarp forest in Thailand". Biodiversity and Conservation. 21 (9): 2287–2298. doi:10.1007/s10531-012-0250-1.
^ Miller OK, Hilton RN (1986). "New and interesting agarics from Western Australia" (PDF). Sydowia. 39: 126–137.
^ van der Westhuizen GC, Eicker A (1987). "Some fungal symbionts of ectotrophic mycorrhizae of pines in South Africa". South African Forestry Journal. 143 (1): 20–24. doi:10.1080/00382167.1987.9630296.
^ Chalermpongse A. (1995). The occurrence and use of ectomycorrhizal fungi in eucalypt plantations in Thailand. In ACIAR PROCEEDINGS (PDF). Canberra: Australian Centre for International Agricultural Research. pp. 127–131.
^ ab Smith SE, Read D (2008). Mycorrhizal Symbiosis (3rd ed.). Amsterdam; Boston: Academic Press. ISBN 978-0-12-370526-6.
^ ab Mühlmann O, Göbl F (June 2006). "Mycorrhiza of the host-specific Lactarius deterrimus on the roots of Picea abies and Arctostaphylos uva-ursi". Mycorrhiza. 16 (4): 245–250. doi:10.1007/s00572-006-0038-9. PMID 16496189.
^ ab Cullings KW, Szaro TM, Bruns TD (1996). "Evolution of extreme specialization within a lineage of ectomycorrhizal epiparasites" (PDF). Nature. 379 (6560): 63–66. doi:10.1038/379063a0.
^ ab Bidartondo MI (August 2005). "The evolutionary ecology of myco-heterotrophy". The New Phytologist. 167 (2): 335–52. doi:10.1111/j.1469-8137.2005.01429.x. PMID 15998389.
^ Yang S, Pfister DH (2006). "Monotropa uniflora plants of eastern Massachusetts form mycorrhizae with a diversity of russulacean fungi". Mycologia. 98 (4): 535–40. doi:10.3852/mycologia.98.4.535. PMID 17139846.
^ Dearnaley JD (September 2007). "Further advances in orchid mycorrhizal research". Mycorrhiza. 17 (6): 475–486. doi:10.1007/s00572-007-0138-1. PMID 17582535.
^ ab Taylor DL, Bruns TD, Hodges SA (January 2004). "Evidence for mycorrhizal races in a cheating orchid". Proceedings. Biological Sciences. 271 (1534): 35–43. doi:10.1098/rspb.2003.2557. PMC 1691555. PMID 15002769.
^ Roy M, Watthana S, Stier A, Richard F, Vessabutr S, Selosse MA (August 2009). "Two mycoheterotrophic orchids from Thailand tropical dipterocarpacean forests associate with a broad diversity of ectomycorrhizal fungi". BMC Biology. 7 (1): 51. doi:10.1186/1741-7007-7-51. PMID 19682351.
^ ab Girlanda M, Selosse MA, Cafasso D, Brilli F, Delfine S, Fabbian R, Ghignone S, Pinelli P, Segreto R, Loreto F, Cozzolino S, Perotto S (February 2006). "Inefficient photosynthesis in the Mediterranean orchid Limodorum abortivum is mirrored by specific association to ectomycorrhizal Russulaceae" (PDF). Molecular Ecology. 15 (2): 491–504. doi:10.1111/j.1365-294x.2005.02770.x. PMID 16448415.
^ Smith ME, Trappe JM, Rizzo DM, Miller SL (May 2006). "Gymnomyces xerophilus sp. nov. (sequestrate Russulaceae), an ectomycorrhizal associate of Quercus in California". Mycological Research. 110 (Pt 5): 575–82. doi:10.1016/j.mycres.2006.03.001. PMID 16769510.
^ Nuytinck J, Verbeken A, Delarue S, Walleyn R (2003). "Systematics of European sequestrate lactarioid Russulaceae with spiny spore ornamentation". Belgian Journal of Botany. 136 (2): 145–153. JSTOR 20794526.
^ Machniki N, Wright LL, Allen A, Robertson CP, Meyer C, Birkebak JM, Ammirati JF (2006). "Russula crassotunicata identified as a host for Dendrocollybia racemosa" (PDF). Pacific Northwest Fungi. 1 (9): 1–7. doi:10.2509/pnwf.2006.001.009.
^ "The Global Fungal Red List Initiative". International Union for Conservation of Nature (IUCN). Retrieved 2014-11-16.
^ "The IUCN Red List of Threatened Species. Version 2014.3". International Union for Conservation of Nature (IUCN). Archived from the original on 2014-06-27. Retrieved 2014-11-15.
^ Evans S, Henrici A, Ing B (2006). "Red Data List of Threatened British Fungi" (PDF). British Mycological Society. Archived from the original (PDF) on 2014-12-14.
^ Senn-Irlet B, Bieri G, Egli S (2007). Rote Liste der gefährdeten Grosspilze der Schweiz [Red list of endangered macromycetes of Switzerland] (PDF) (in German). Bern; Birmensdorf: Bundesamt für Umwelt & WSL.
^ Holec J, Beran M (2006). "Red list of fungi (macromycetes) of the Czech Republic" (PDF). Příroda (in Czech and English). 24: 1–282. ISSN 1211-3603.
^ "Nationally Critical: Rare and endangered fungi". Landcare Research. Retrieved 2014-11-15.
^ Parkyn L, Stoneman RE, Ingram HA, eds. (1997). Conserving peatlands. Wallingford, UK: CAB International. ISBN 978-0-85198-998-3.
^ Médail F, Quézel P (1999). "Biodiversity Hotspots in the Mediterranean Basin: Setting global conservation priorities". Conservation Biology. 13 (6): 1510–1513. doi:10.1046/j.1523-1739.1999.98467.x.
^ Sjumpangani S, Chirwa PW, Akinnifesi FK, Sileshi G, Ajayi O (2009). "The miombo woodlands at the cross roads: Potential threats, sustainable livelihoods, policy gaps and challenges". Natural Resources Forum. 33 (2): 150–159. doi:10.1111/j.1477-8947.2009.01218.x.
^ Jonsson BG, Kruys N, Ranius T (2005). "Ecology of species living on dead wood – Lessons for dead wood management" (PDF). Silva Fennica. 39 (2): 289–309. doi:10.14214/sf.390.
^ Bickford D, Lohman DJ, Sodhi NS, Ng PK, Meier R, Winker K, Ingram KK, Das I (March 2007). "Cryptic species as a window on diversity and conservation". Trends in Ecology & Evolution. 22 (3): 148–55. doi:10.1016/j.tree.2006.11.004. PMID 17129636.
^ "Russulales News / Edibility and toxicity of Russulales". Museo delle scienze. Retrieved 2014-12-13.
^ Zeitlmayr L (1976). Wild Mushrooms: An Illustrated Handbook. London, UK: Transatlantic Arts. p. 44. ISBN 978-0-584-10324-3.
^ Arora D (1986). Mushrooms Demystified: A Comprehensive Guide to the Fleshy Fungi. Berkeley, California: Ten Speed Press. p. 84. ISBN 978-0-89815-169-5.
^ Miller HR, Miller OK Jr (2006). North American Mushrooms: A Field Guide to Edible and Inedible Fungi. Guilford, Connecticut: Falcon Guides. p. 73. ISBN 978-0-7627-3109-1.
^ Veteläinen M, Huldén M, Pehu T (2008). State of plant genetic resources for food and agriculture in Finland. Second Finnish National Report (PDF). Country Report on the State of Plant Genetic Resources for Food and Agriculture (Report). Sastamala, Finland: Ministry of Agriculture and Forestry. p. 14.
^ Molokhovets E (1992). Classic Russian Cooking. Bloomington, Indiana: Indiana University Press. p. 95. ISBN 978-0-253-21210-8.
^ Roberts P, Evans S (2014). The Book of Fungi: A Life-Size Guide to Six Hundred Species from around the World. Chicago: University of Chicago Press. p. 595. ISBN 978-0-226-17719-9.
^ Rogers R (2006). The Fungal Pharmacy: Medicinal Mushrooms of Western Canada. Edmonton, Alberta: Prairie Deva Press. p. 178. ISBN 978-0-9781358-1-2.
^ Chen Z, Zhang P, Zhang Z (2014). "Investigation and analysis of 102 mushroom poisoning cases in Southern China from 1994 to 2012". Fungal Diversity. 64 (1): 123–131. doi:10.1007/s13225-013-0260-7.
^ ab Suortti T, von Wright A, Koskinen A (1983). "Necatorin, a highly mutagenic compound from Lactarius necator". Phytochemistry. 22 (12): 2873–2874. doi:10.1016/S0031-9422(00)97723-9.
^ Guerin-Laguette A, Cummings N, Butler RC, Willows A, Hesom-Williams N, Li S, Wang Y (October 2014). "Lactarius deliciosus and Pinus radiata in New Zealand: towards the development of innovative gourmet mushroom orchards". Mycorrhiza. 24 (7): 511–23. doi:10.1007/s00572-014-0570-y. PMID 24676792.
^ Rapior S, Fons F, Bessière JM (2000). "The fenugreek odor of Lactarius helvus". Mycologia. 92 (2): 305–308. doi:10.2307/3761565. JSTOR 3761565.
^ Wood WF, Brandes JA, Foy BD, Morgan CG, Mann TD, DeShazer DA (2012). "The maple syrup odour of the "candy cap" mushroom, Lactarius fragilis var. rubidus". Biochemical Systematics and Ecology. 43: 51–53. doi:10.1016/j.bse.2012.02.027.
^ Harmon AD, Weisgraber KH, Weiss U (1979). "Preformed azulene pigments of Lactarius indigo (Schw.) Fries (Russulaceae, Basidiomycetes)". Cellular and Molecular Life Sciences. 36 (1): 54–56. doi:10.1007/BF02003967.
^ Fröde R, Bröckelmann M, Steffan B, Steglich W, Marumoto R (1995). "A novel type of triterpenoid quinone methide pigment from the toadstool Russula flavida (Agaricales)". Tetrahedron. 51 (9): 2553–2560. doi:10.1016/0040-4020(95)00012-W.
^ Gry J, Andersson C (2014). Mushrooms Traded as Food. Vol II sec. 2. Copenhagen: Nordic Council of Ministers. p. 236. ISBN 978-92-893-2705-3.
^ abcdefg Vitari G, Vita-Finzi P (1995). "Sesquiterpenes and other secondary metabolites of genus Lactarius (Basidiomycetes): Chemistry and biological activity". Studies in Natural Products Chemistry. Structure and Chemistry (Part D). Elsevier Science. 17: 153. ISBN 978-0-08-054198-3.
^ Kobata K, Kano S, Shibata H (1995). "New lactarane sesquiterpenoid from the fungus Russula emetica". Bioscience, Biotechnology, and Biochemistry. 59 (2): 316–318. doi:10.1271/bbb.59.316.
^ Tanaka Y, Kawahara S, Eng AH, Takei A, Ohya N (1994). "Structure of cis-polyisoprene from Lactarius mushrooms". Acta Biochimica Polonica. 41 (3): 303–9. PMID 7856401.
^ Yue JM, Chen SN, Lin ZW, Sun HD (April 2001). "Sterols from the fungus Lactarium volemus". Phytochemistry. 56 (8): 801–6. doi:10.1016/S0031-9422(00)00490-8. PMID 11324907.
^ Bourquelot E (1889). "Sur la volémite, nouvelle matière sucrée" [On volémite, a new sweet substance]. Journal de Pharmacie et de Chimie (in French). 2: 385–390.
^ Matsuura M, Saikawa Y, Inui K, Nakae K, Igarashi M, Hashimoto K, Nakata M (July 2009). "Identification of the toxic trigger in mushroom poisoning". Nature Chemical Biology. 5 (7): 465–7. doi:10.1038/nchembio.179. PMID 19465932.
^ Yaltirak T, Aslim B, Ozturk S, Alli H (August 2009). "Antimicrobial and antioxidant activities of Russula delica Fr". Food and Chemical Toxicology. 47 (8): 2052–6. doi:10.1016/j.fct.2009.05.029. PMID 19481130.
^ Zhang G, Sun J, Wang H, Ng TB (August 2010). "First isolation and characterization of a novel lectin with potent antitumor activity from a Russula mushroom". Phytomedicine. 17 (10): 775–81. doi:10.1016/j.phymed.2010.02.001. PMID 20378319.
External links
Wikispecies has information related to Russulaceae |
- Russulales News: Database, literature, identification
- Peat Bog and Mire Russulaceae
- Ghent University, Mycology: An integrated approach to the phylogeny and phylogeography of the Russulaceae (Basidiomycota)


