Jumping spider
| Jumping spiders | |
|---|---|
 | |
| An adult male Phidippus audax | |
Scientific classification | |
| Kingdom: | Animalia |
| Phylum: | Arthropoda |
| Subphylum: | Chelicerata |
| Class: | Arachnida |
| Order: | Araneae |
| Infraorder: | Araneomorphae |
| Family: | Salticidae Blackwall, 1841 |
| Genera | |
See List of Salticidae genera. | |
Diversity | |
600+ genera, 5000+ species | |
 | |
Jumping spiders are a group of spiders that constitute the family Salticidae. This family contains roughly 635 described genera and about 6080 described species,[1] making it the largest family of spiders with about 13% of all species.[2] Jumping spiders have some of the best vision among arthropods and use it in courtship, hunting, and navigation. Although they normally move unobtrusively and fairly slowly, most species are capable of very agile jumps, notably when hunting, but sometimes in response to sudden threats or crossing long gaps. Both their book lungs and tracheal system are well-developed, and they use both systems (bimodal breathing). Jumping spiders are generally recognized by their eye pattern. All jumping spiders have four pairs of eyes, with the anterior median pair being particularly large.
Contents
1 Distinguishing characteristics
2 Habitat
3 Vision
4 Behavior
4.1 Hunting
5 Diet
6 Reproduction
6.1 General mating behavior
6.2 Visual and non-visual courtship displays
6.3 Consequences of sexual dimorphism
7 Taxonomy
8 Models for mimicry
9 Fossils
10 See also
11 References
12 Further reading
13 External links
Distinguishing characteristics

Salticidae male anterior and dorsal aspects, showing positions of eyes
Jumping spiders are among the easiest to distinguish from similar spider families because of the shape of the cephalothorax and their eye patterns. The families closest to Salticidae in general appearance are the Corinnidae (distinguished also by prominent spines on the back four legs), the Oxyopidae (the lynx spiders, distinguished by very prominent spines on all legs), and the Thomisidae (the crab spiders, distinguished by their front four legs, which are very long and powerful). None of these families however, has eyes that resemble those of the Salticidae. Conversely, the legs of jumping spiders are not covered with any very prominent spines. Their front four legs generally are larger than the hind four, but not as dramatically so as those of the crab spiders, nor are they held in the outstretched-arms attitude characteristic of the Thomisidae.[3] In spite of the length of their front legs, Salticidae depend on their rear legs for jumping. The generally larger front legs are used partly to assist in grasping prey,[4] and in some species, the front legs and pedipalps are used in species-recognition signalling.
The jumping spiders, unlike the other families, have faces that are roughly rectangular surfaces perpendicular to their direction of motion. In effect this means that their forward-looking, anterior eyes are on "flat faces", as shown in the photographs. Their eye pattern is the clearest single identifying characteristic. They have eight eyes, as illustrated.[3][4] Most diagnostic are the front row of four eyes, in which the anterior median pair are more dramatically prominent than any other spider eyes apart from the posterior median eyes of the Deinopidae. There is, however, a radical functional difference between the major (AME) eyes of Salticidae and the major (PME) eyes of the Deinopidae; the large posterior eyes of Deinopidae are adapted mainly to vision in dim light, whereas the large anterior eyes of Salticidae are adapted to detailed, three-dimensional vision for purposes of estimating the range, direction, and nature of potential prey, permitting the spider to direct its attacking leaps with great precision. The anterior lateral eyes, though large, are smaller than the AME and provide a wider forward field of vision.
The rear row of four eyes may be described as strongly bent, or as being rearranged into two rows, with two large posterior lateral eyes furthest back. They serve for lateral vision. The posterior median eyes also have been shifted out laterally, almost as far as the posterior lateral eyes. They are usually much smaller than the posterior lateral eyes and there is doubt about whether they are at all functional in many species.
The body length of jumping spiders generally range from 1 to 25 mm (0.04–0.98 in).[3][5] The largest is Hyllus giganteus,[5] while other genera with relatively large species include Phidippus, Philaeus and Plexippus.[6]
In addition to using their silk for safety lines while jumping, they also build silken "pup tents", where they shelter from bad weather and sleep at night. They molt within these shelters, build and store egg cases within them, and also spend the winter in them.[7]
Habitat
Jumping spiders live in a variety of habitats. Tropical forests harbor the most species, but they are also found in temperate forests, scrub lands, deserts, intertidal zones, and mountainous regions. Euophrys omnisuperstes is the species reported to have been collected at the highest elevation, on the slopes of Mount Everest.[8]
Vision
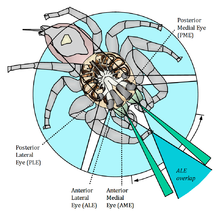
The visual fields of a jumping spider

The eight eyes of a Telamonia dimidiata located near the front

Marpissa muscosa, female
Jumping spiders have four pairs of eyes; three secondary pairs that are fixed and a principal pair that is movable.
The posterior median eyes (PME) are vestigial in many species, but in some primitive sub-families they are comparable in size with the other secondary eyes and help to detect motion.[9] While unable to form images, it is suspected that the reduced pair of eyes has a role similar to that of insect ocelli by receiving light from the sky. The photoreceptors in the other secondary pairs are almost exclusively greensensitive, but the PME have two visual pigments different from that in all the other eyes, sensitive to blue and UV-light.[10]
The posterior lateral eyes (PLE) are wide-angle motion detectors which sense motions from the side and behind. Combined with the other eyes, it gives the spider a near 360-degree view of the world.
The anterior lateral eyes (ALE) have the best visual acuity of the secondary eyes.[11] It has been shown that they are able to distinguish some details as well, and without them no "looming response" will be triggered by motion.[12] Even with all the other pairs covered, jumping spiders in a study could still detect, stalk and attack flies, using the anterior lateral eyes only, which are also sufficiently widely spaced to provide stereoscopic vision.[13]
The anterior median eyes (AME) have very good vision. This pair of eyes is built like a telescopic tube with a corneal lens in the front and a second lens in the back that focus images onto a four-layered retina, a narrow boomerang-shaped strip oriented vertically.[14][15] Physiological experiments have shown they may have up to four different kinds of receptor cells, with different absorption spectra, giving them the possibility of tetrachromatic color vision, with sensitivity extending into the ultraviolet range.[16] As the eyes are too close together to allow depth perception, and the animals do not make use of motion parallax, they have evolved a method called image defocus instead. Of the four photoreceptor layers in the retina, the first two closest to the surface contain ultraviolet-sensitive pigments while the two deepest contain green-sensitive pigments. The incoming green light is only focused on the deepest layer, while the other one receives defocused or fuzzy images. By measuring the amount of defocus from the fuzzy layer, it is possible to calculate the distance to the objects in front of them.[17][18] In addition to receptor cells, also red filters have been detected, located in front of the cells that normally register green light.[19] It seems that all salticids, regardless of whether they have two, three, or four kinds of color receptors, are highly sensitive to UV light.[16] Some species (for example, Cosmophasis umbratica) are highly dimorphic in the UV spectrum, suggesting a role in sexual signaling (Lim & Li, 2005). Color discrimination has been demonstrated in behavioral experiments.
The principal, anterior median, eyes have high resolution (11 min visual angle),[20] but the field of vision is narrow, from 2 to 5°. The central region of the retina, where acuity is highest, is no more than six or seven receptor rows wide. However, the eye can scan objects off the direct axis of vision. As the lens is attached to the carapace, the eye's scanning movements are restricted to its retina through a complicated pattern of translations and rotations.[21] This dynamic adjustment is a means of compensation for the narrowness of the static field of vision. It is analogous to the way most primates move their eyes to focus images of interest onto the fovea centralis. Such movements within the jumping spider's eyes are visible from outside when the attention of the spider is directed to various targets.[22]
Behavior
Jumping spiders are generally diurnal, active hunters. Their well-developed internal hydraulic system extends their limbs by altering the pressure of body fluid (hemolymph) within them. This enables the spiders to jump without having large muscular legs like a grasshopper. Most jumping spiders can jump several times the length of their bodies. When a jumping spider is moving from place to place, and especially just before it jumps, it tethers a filament of silk (or 'dragline') to whatever it is standing on to protect itself if the jump should fail.[7] Should it fall, for example if the prey shakes it off, it climbs back up the silk tether. Some species, such as Portia, will actually let themselves down to attack prey such as a web spider apparently secure in the middle of its web. Like many other spiders that leave practically continuous silk trails, jumping spiders impregnate the silk line with pheromones that play a role in social and reproductive communication, and possibly in navigation.
Certain species of jumping spiders have been shown by experiment to be capable of learning, recognizing, and remembering colors, and adapting their hunting behavior accordingly.[23]
Hunting
The hunting behaviour of the Salticidae is confusingly varied compared to that of most spiders in other families.[24] Salticids hunt diurnally as a rule, which is consistent with their highly developed visual system. When it detects potential prey, a jumping spider typically begins orienting itself by swivelling its cephalothorax to bring the anterior median eyes to bear. It then moves its abdomen into line with its cephalothorax. After that, it might spend some time inspecting the object of its attention and determining whether a camouflaged or doubtful item of prey is promising, before it starts to stalk slowly forward. When close enough, the spider pauses to attach a dragline, then springs onto the prey.
There are, though, many variations on the theme and many surprising aspects. For one thing, salticids do not necessarily follow a straight path in approaching prey. They may follow a circuitous course, sometimes even a course that takes the hunter through regions from which the prey is not visible. Such complex adaptive behaviour is hard to reconcile with an organism that has such a tiny brain, but some jumping spiders, in particular some species of Portia, can negotiate long detours from one bush down to the ground, then up the stem of another bush to capture a prey item on a particular leaf. Such behaviour still is the subject of research.[24]
Some salticid species are continually on the move, stopping periodically to look around for prey, which they then stalk immediately. Others spend more time scanning their surroundings from one position, actively stalking any prey they detect. Members of the genus Phaeacius take that strategy to extremes; they sit on a tree trunk, facing downwards and rarely do any stalking, but simply lunge down on any prey items that pass close before them.[24]
Some Salticidae specialise in particular classes of prey. Ants comprise one such class. Most spiders, including most salticids, avoid worker ants, but several species not only eat them as a primary item in their diets, but also employ specialised attack techniques — Corythalia canosa for example, circles round to the front of the ant and grabs it over the back of its head. Such myrmecophagous species, however, will not necessarily refuse other prey items, and will routinely catch flies and similar prey in the usual salticid fashion, without the special precautions they apply in hunting dangerous prey such as ants. Ants offer the advantages of being plentiful prey items for which there is little competition from other predators, but it remains profitable to catch less hazardous prey when it presents itself.[24]
Some of the most surprising hunting behaviour occurs among the araneophagous Salticidae, and it varies greatly in method. Many of the spider-hunting species quite commonly will attack other spiders, whether fellow salticids or not, in the same way as any other prey, but some kinds resort to web invasion; nonspecialists such as Phidippus audax sometimes attack prey ensnared in webs, basically in acts of kleptoparasitism — sometimes they leap onto and eat the web occupant itself, or simply walk over the web for that purpose.
Salticidae in the genera Brettus, Cyrba, Gelotia, and Portia display more advanced web-invasion behavior. They slowly advance onto the web and vibrate the silk with their pedipalps and legs. In this respect, their behaviour resembles that of the Mimetidae, probably the most specialised of the araneophagous spider families. If the web occupant approaches in the manner appropriate to dealing with ensnared prey, the predator attacks.[24]
The foregoing examples present the Salticidae as textbook examples of active hunters; they would hardly seem likely to build webs other than those used in reproductive activities, and in fact, most species really do not build webs to catch prey. However, exceptions occur, though even those that do build capture webs generally also go hunting like other salticids. Some Portia species, for example, spin capture webs that are functional, though not as impressive as some orb webs of the Araneidae; Portia webs are of an unusual funnel shape and apparently adapted to the capture of other spiders. Spartaeus species, on the other hand, largely capture moths in their webs. In their review of the ethology of Salticidae, Richman and Jackson speculate on whether such web building is a relic of the evolution of this family from web-building ancestors.[24]
In hunting, Salticidae also use their silk for a tether to enable them to reach prey that otherwise would be inaccessible. For example, by advancing towards the prey to less than the jumping distance, then retreating and leaping in an arc at the end of the tether line, many species can leap onto prey on vertical or even on inverted surfaces, which of course in a gravitational field would not be possible without such a tether.
Having made contact with the prey, hunting Salticidae administer a bite to inject rapidly acting venom that gives the victim little time to react.[25] In this respect, they resemble the Mimetidae and Thomisidae, families that ambush prey that often are larger than the predator, and they do so without securing the victim with silk; they accordingly must immobilise it immediately and their venom is adapted accordingly.
@media all and (max-width:720px){.mw-parser-output .tmulti>.thumbinner{width:100%!important;max-width:none!important}.mw-parser-output .tmulti .tsingle{float:none!important;max-width:none!important;width:100%!important;text-align:center}}



Diet

A camouflaged Menemerus sp. jumping spider with a captured male ant
Although jumping spiders are generally carnivorous, many species have been known to include nectar in their diets,[26] and one species, Bagheera kiplingi, feeds primarily on plant matter.[27] None are known to feed on seeds or fruit. Extrafloral nectaries on plants, such as Chamaecrista fasciculata (partridge pea), provide jumping spiders with nectar; the plant benefits accordingly when the spiders prey on whatever pests they find.
Reproduction
General mating behavior
Jumping spiders conduct complex, visual courtship displays using both movements and physical bodily attributes. Unlike females, males possess plumose hairs, colored or iridescent hairs (particularly pronounced in the peacock spiders), front leg fringes, structures on other legs, and other, often bizarre, modifications. These characteristics are used in a courtship "dance" in which the colored or iridescent parts of the body are displayed. In addition to the display of hairs, jumping spiders perform complex sideling, vibrational, or zigzag movements to attract females. If receptive to the male, the female will assume a passive, crouching position. It has also recently been discovered that many males have auditory signals as well. These amplified sounds presented to the females resemble buzzes or drum rolls.[28] In some species, the female may also vibrate her palps or abdomen. The male will then extend his front legs towards the female to touch her. If the female remains receptive, the male will climb on the female's back and inseminate her with his palps.[29]
Visual and non-visual courtship displays

Courtship display of Saitis barbipes jumping spider
Species vary greatly in visual and vibratory components of courtship.[30] Many species have patches of UV reflectance, which are exhibited in mature males.[31] This visual component is used by some female jumping spiders for mate choice.[32] Body size dimorphism seems to play a role in male-male competition, where the larger bodied males will have a fitness advantage to hook the competitor and push down its legs into a vulnerable state, thus impairing the competitor's ability to fight back.[33] Since larger spiders have a better chance of winning the fight, they also have a greater chance to successfully mate and will be favored by sexual selection.[34]
Consequences of sexual dimorphism
Maintaining UV ornamentation and size dimorphism of some jumping spiders may seem strictly beneficial to sexual selection, yet there are costs to maintain such distinguishing characteristics.[32] While UV reflecting individuals may attract more female spiders, it can also increase the risk of predation.[15] Similarly, larger body size can be costly, as it may also increase the risk of predation.[15]
Taxonomy
| Classification within the spiders (Araneae)[35] | ||||||||||||||||||||||||||||||||||||||||||||||||||||||||||||
.mw-parser-output table.clade{border-spacing:0;margin:0;font-size:100%;line-height:100%;border-collapse:separate;width:auto}.mw-parser-output table.clade table.clade{width:100%}.mw-parser-output table.clade td{border:0;padding:0;vertical-align:middle;text-align:center}.mw-parser-output table.clade td.clade-label{width:0.8em;border:0;padding:0 0.2em;vertical-align:bottom;text-align:center}.mw-parser-output table.clade td.clade-slabel{border:0;padding:0 0.2em;vertical-align:top;text-align:center}.mw-parser-output table.clade td.clade-bar{vertical-align:middle;text-align:left;padding:0 0.5em}.mw-parser-output table.clade td.clade-leaf{border:0;padding:0;text-align:left;vertical-align:middle}.mw-parser-output table.clade td.clade-leafR{border:0;padding:0;text-align:right}
|
The monophyly of the family Salticidae is well established through both phylogenetic and morphological analyses, but no consensus exists on what other group of spiders are most closely related to the jumping spiders. Suggested sister groups have included the oxyopids (lynx spiders), thomisids (crab spiders), clubionoids (sac spiders), and web-building spiders.[36] A 2017 phylogenetic analysis suggests that the sister group may be the family Miturgidae.[37]
The taxonomy within the jumping spider family was significantly revised in 2015. The family is now divided into seven subfamilies:[1]
Onomastinae Maddison, 2015 – 1 genus, 12 species
Asemoneinae Maddison, 2015 – 5 genera, 38 species
Lyssomaninae Blackwall, 1877 – 2 genera, 92 species
Spartaeinae Wanless, 1984 – 29 genera, 165 species
Eupoinae Maddison, 2015 – 3 genera, 34 species
Hisponinae Simon, 1901 – 9 genera, 53 species
Salticinae Blackwall, 1841 – 538 genera, 5379 species
Of these subfamilies, Salticinae is by far the largest, comprising over 90% of the known species of jumping spiders.[1]
Models for mimicry
Some small insects are thought to have evolved an appearance or behavioural traits that resemble those of jumping spiders and this is suspected to prevent their predation, specifically from jumping spiders. Some examples appear to be provided by patterns on the wings of some tephritid flies,[38][39] nymph of a fulgorid[40] and possibly some moths.[41]
Fossils
Very few jumping spider fossils have been found. Of those known, all are from Cenozoic era amber. The oldest fossils are from Baltic amber dating to the Eocene epoch, specifically, 54 to 42 million years ago. Other fossil jumping spiders have been found in Chiapan amber and Dominican amber.[42]
See also
List of Salticidae genera – List of Salticidae species
Peckhamia (journal)- Spider taxonomy
References
^ abc Maddison, Wayne P. (November 2015). "A phylogenetic classification of jumping spiders (Araneae: Salticidae)". Journal of Arachnology. 43 (3): 231–292. doi:10.1636/arac-43-03-231-292..mw-parser-output cite.citation{font-style:inherit}.mw-parser-output q{quotes:"""""""'""'"}.mw-parser-output code.cs1-code{color:inherit;background:inherit;border:inherit;padding:inherit}.mw-parser-output .cs1-lock-free a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/6/65/Lock-green.svg/9px-Lock-green.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-limited a,.mw-parser-output .cs1-lock-registration a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/d/d6/Lock-gray-alt-2.svg/9px-Lock-gray-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-lock-subscription a{background:url("//upload.wikimedia.org/wikipedia/commons/thumb/a/aa/Lock-red-alt-2.svg/9px-Lock-red-alt-2.svg.png")no-repeat;background-position:right .1em center}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration{color:#555}.mw-parser-output .cs1-subscription span,.mw-parser-output .cs1-registration span{border-bottom:1px dotted;cursor:help}.mw-parser-output .cs1-hidden-error{display:none;font-size:100%}.mw-parser-output .cs1-visible-error{font-size:100%}.mw-parser-output .cs1-subscription,.mw-parser-output .cs1-registration,.mw-parser-output .cs1-format{font-size:95%}.mw-parser-output .cs1-kern-left,.mw-parser-output .cs1-kern-wl-left{padding-left:0.2em}.mw-parser-output .cs1-kern-right,.mw-parser-output .cs1-kern-wl-right{padding-right:0.2em}
^ Peng, Xian-Jin; Tso, I-Min & Li, Shu-Qiang (2002). "Five New and Four Newly Recorded Species of Jumping Spiders from Taiwan (Araneae: Salticidae)" (PDF). Zoological Studies. 41 (1): 1–12. Retrieved 28 January 2016.
^ abc Richman, D.B.; Edwards, G.B. & Cutler, B. (2005). "Salticidae". In Ubick, D.; Paquin, P.; Cushing, P.E. & Roth, V. Spiders of North America: an identification manual. American Arachnological Society. pp. 205–216. ISBN 978-0-9771439-0-0.
^ ab Crompton, J. (1954). The Life of the Spider. New York: New American Library (reprint). p. 77. OCLC 2896911.
^ ab "Watch the world's biggest jumping spider make a leap". BBC Earth. 2016-01-29. Retrieved 2016-06-18.
^ Macík, Stanislav (2012-08-27). "Phiddipus regius: the Jewel between Spider Predators". arachnos.eu. Retrieved 2016-06-18.
^ ab Foelix, Rainer F. (1996). Biology of Spiders. Oxford University Press. p. 11. ISBN 978-0-674-07431-6.
^ Wanless, F. R. (1975). "Spiders of the family Salticidae from the upper slopes of Everest and Makalu". Bulletin of the British Arachnological Society. 3 (5): 132–136.
^ "short communication fields of view of the eyes – The Company of Biologists Limited 1985" (PDF). Retrieved 13 August 2013.
^ Functional Properties of Opsins and their Contribution to Light-Sensing Physiology
^ Zurek, Daniel B.; Nelson, Ximena J. (August 2012). "Hyperacute motion detection by the lateral eyes of jumping spiders". Vision Research. 66: 26–30. doi:10.1016/j.visres.2012.06.011. PMID 22750020.
^ "Jeepers, Peepers: Why Spiders Have So Many Eyes". Livescience.com. 17 October 2012. Retrieved 13 August 2013.
^ Zurek, D. B.; Taylor, A. J.; Evans, C. S.; Nelson, X. J. (25 June 2010). "The role of the anterior lateral eyes in the vision-based behaviour of jumping spiders". Journal of Experimental Biology. 213 (14): 2372–2378. doi:10.1242/jeb.042382. PMID 20581266.
^ "Eye on the Web". Archopht.jamanetwork.com. 21 August 2007. Retrieved 13 August 2013.
^ abc Harland, D.P. & Jackson, R.R. (2000). "'Eight-legged cats' and how they see – a review of recent research on jumping spiders (Araneae: Salticidae)". Cimbebasia. 16: 231–240. Retrieved 28 January 2016.
^ ab Peaslee, A.G. & Wilson, G. (May 1989). "Spectral sensitivity in jumping spiders (Araneae, Salticidae)". Journal of Comparative Physiology A. 164 (3): 359–63. doi:10.1007/BF00612995. PMID 2709341.
^ "Jumping Spiders' Unique Vision Revealed". Livescience.com. 26 January 2012. Retrieved 13 August 2013.
^ Nagata, T.; et al. (2012). "Depth Perception from Image Defocus in a Jumping Spider". Science. 335 (6067): 469–71. doi:10.1126/science.1211667. PMID 22282813.
^ Filters let jumping spiders spot flashy mates
^ Land, MF (1969). "Structure of the Retinae of the Principal Eyes of Jumping Spiders (Salticidae: Dendryphantinae) in Relation to Visual Optics". The Journal of Experimental Biology. 51 (2): 443–70. PMID 5351425.
^ "Topic: Scanning eyes in molluscs and arthropods". Mapoflife.org. Retrieved 13 August 2013.
^ Land, M. F. (1969). "Movements of the retinae of jumping spiders (Salticidae: Dendryphantinae) in response to visual stimuli" (PDF). The Journal of Experimental Biology. 51 (2): 471–93. PMID 5351426.
^ Jakob, Elizabeth M.; et al. (2007). "Jumping spiders associate food with color cues in a T-maze" (PDF). Journal of Arachnology. 35 (3): 487–492. doi:10.1636/JOA-ST06-61.1.
^ abcdef Richman, David B.; Jackson, Robert R. (1992). "A review of the ethology of jumping spiders (Araneae, Salticidae)" (PDF). Bull. Br. Arachnol. Soc. 9 (2): 33–37.
^ National Geographic video of capture of bee by jumping spider. Youtube.com (27 February 2009). Retrieved on 4 May 2013.
^ Jackson, Robert R.; Simon D. Pollard; Ximena J. Nelson; G. B. Edwards; Alberto T. Barrion (2001). "Jumping spiders (Araneae: Salticidae) that feed on nectar" (PDF). Journal of Zoology, London. 255: 25–29. doi:10.1017/S095283690100108X.
^ Milius, Susan (30 August 2008). "Vegetarian Spider". Science News. Retrieved 9 April 2009.
^ Elias, DO; Mason, AC; Maddison, WP; Hoy, RR (2003). "Seismic signals in a courting male jumping spider". The Journal of Experimental Biology. 206 (22): 4029–4039. doi:10.1242/jeb.00634. PMID 14555743.
^ Foelix, Rainer F. (1996). Biology of Spiders. Oxford University Press. pp. 195–197. ISBN 978-0-674-07431-6.
^ Morelle, Rebecca (2 May 2008) " Study sheds light on spider sex", BBC News.
^ (Lim, Matthew L. M., and
Daiqin Li. "Courtship and Male-Male Agonistic Behaviour of Comsophasis
Umbratica Simon, an Ornate Jumping Spider (Araneae: Salticidae)." The Raffles
Bulletin of Zoology (2004): 52(2): 435-448. National University of Singapore.
Web. 20 September 2015.)
^ ab Bulbert, Matthew W., James C. O’Hanlon, Shane
Zappettini, Shichang Zhang, and Daiqin Li. "Sexually Selected UV Signals in the
Tropical Ornate Jumping Spider, Cosmophasis umbratica, May Incur Costs from
Predation." Ecology and Evolution (2015): 5(4): 914-920. John Wiley & Sons
Ltd. Web. 20 September 2015.
^ Lim, Matthew L. M.; Li, Daiqin (2006). "Extreme Ultraviolet Sexual Dimorphism in Jumping Spiders (Araneae: Salticidae)". Biological Journal of the Linnean Society. 89 (3): 397–406. doi:10.1111/j.1095-8312.2006.00704.x.
^ Lim, Matthew L. M.; Li, Daiqin (2004). "Courtship and Male-Male Agonistic Behaviour of Cosmophasis umbratica Simon, an Ornate Jumping Spider (Araneae: Salticidae)". The Raffles Bulletin of Zoology. 52 (2): 435–448.
^ Wheeler, Ward C.; Coddington, Jonathan A.; Crowley, Louise M.; et al. (December 2016). "The spider tree of life: phylogeny of Araneae based on target-gene analyses from an extensive taxon sampling". Cladistics. 33 (6): 574–616. doi:10.1111/cla.12182.
^ Maddison, Wayne P.; Hedin, Marshal C. (2003). "Jumping spider phylogeny (Araneae:Salticidae)". Invertebrate Systematics. 17 (4): 529–549. doi:10.1071/IS02044.
^ Maddison, Wayne P.; Evans, Samuel C.; Hamilton, Chris A.; Bond, Jason E.; Lemmon, Alan R.; Lemmon, Emily Moriarty (4 September 2017). "A genome-wide phylogeny of jumping spiders (Araneae, Salticidae), using anchored hybrid enrichment". ZooKeys. 695 (695): 89–101. doi:10.3897/zookeys.695.13852. PMC 5673835. PMID 29134008.
^
Whitman, D.W, Orsak L & Greene E. (1988). "Spider mimicry in fruit flies (Diptera: Tephritidae): Further experiments on the deterrence of jumping spiders (Araneae: Salticidae) by Zonosemata vittigera (Coquillett)". Annals of the Entomological Society of America. 81 (3): 532–536. doi:10.1093/aesa/81.3.532.CS1 maint: Multiple names: authors list (link)
^ Rao, D.; Díaz-Fleischer, F. (2012). "Characterisation of Predator-Directed Displays in Tephritid Flies". Ethology. 118 (12): 1165–1172. doi:10.1111/eth.12021.
^
Zolnerowich, Gregory (1992). "A Unique Amycle Nymph (Homoptera: Fulgoridae) That Mimics Jumping Spiders (Araneae: Salticidae)". Journal of the New York Entomological Society. 100 (3): 498–502. JSTOR 25009980.
^ Rota J, Wagner DL (2006). "Predator Mimicry: Metalmark Moths Mimic Their Jumping Spider Predators". PLoS ONE. 1 (1): e45. doi:10.1371/journal.pone.0000045. PMC 1762363. PMID 17183674.
^ Hill, David Edwin (7 October 2009). "Salticidae of the Antarctic land bridge" (PDF). Peckhamia.
Further reading
.mw-parser-output .refbegin{font-size:90%;margin-bottom:0.5em}.mw-parser-output .refbegin-hanging-indents>ul{list-style-type:none;margin-left:0}.mw-parser-output .refbegin-hanging-indents>ul>li,.mw-parser-output .refbegin-hanging-indents>dl>dd{margin-left:0;padding-left:3.2em;text-indent:-3.2em;list-style:none}.mw-parser-output .refbegin-100{font-size:100%}
Vasilevsky, M. (2012). A Classical Taxonomic Guide to Identifying Fifty Unique North American Jumping Spiders. Lulu.
[self-published source?]
Kaston, B.J. (1953). How to Know the Spiders. Pictured key nature series (1st ed.). Dubuque, IA: W.C. Brown Co. OCLC 681432632.
Forster, L.M. (1982). "Vision and prey-catching strategies in jumping spiders". American Scientist. 70: 165–175.
Jackson, R.R. (1982). "The behavior of communicating in jumping spiders (Salticidae)". In Witt, P.; Rovner, J. Spider Communication Mechanisms and Ecological Significance. Princeton, NJ: Princeton University Press. pp. 213–247. OCLC 951407473.
Jackman, John A. (1997). A Field Guide to Spiders & Scorpions of Texas. Houston, TX: Gulf Publishing Company. p. 127.
Nakamura, T.; Yamashita, S. (2000). "Learning and discrimination of colored papers in jumping spiders (Araneae, Salticidae)". Journal of Comparative Physiology A. 186 (9): 897–901. doi:10.1007/s003590000143.
Elias, D.O.; Mason, A.C.; Maddison, W.P.; Hoy, R.R. (2003). "Seismic signals in a courting male jumping spider (Araneae: Salticidae)". Journal of Experimental Biology. 206 (22): 4029–4039. doi:10.1242/jeb.00634. PMID 14555743.
Lim, M.L.M.; Li, D. (2005). "Extreme ultraviolet sexual dimorphism in jumping spiders (Araneae: Salticidae)". Biological Journal of the Linnean Society. 89 (3): 397–406. doi:10.1111/j.1095-8312.2006.00704.x.
External links
| Wikimedia Commons has media related to Salticidae. |
Wikispecies has information related to Salticidae |
- Asian jumping spiders and photo references
- Comprehensive resource on the morphology and taxonomy of jumping spiders (Salticidae): www.jumping-spiders.com
- Global Species Database of Salticidae
- Video of a jumping spider's mating behavior
- World Spider Catalog
- Jumping Spiders of NW-Europe
- Jumping spiders of Australia
- American Jumping Spiders – 70 Species Videos (includes introduction to salticids, predation, mating, and other behaviors)
- Movies of Habronattus courtship behavior
- Male jumping spider courtship dance with contact microphone picking up and amplifying sounds
- The Australian Faunal Directory taxonomic classification of Australian salticids


